The latest papers from Ning et al. Cell (2019) and Anthony JIES (2019) have offered some interesting new data, supporting once more what could be inferred since 2015, and what was evident in population genomics since 2017: that Proto-Indo-Europeans expanded under R1b bottlenecks, and that the so-called “Steppe ancestry” referred to two different components, one – Yamnaya or Steppe_EMBA ancestry – expanding with Proto-Indo-Europeans, and the other one – Corded Ware or Steppe_MLBA ancestry – expanding with Uralic speakers.
The following maps are based on formal stats published in the papers and supplementary materials from 2015 until today, mainly on Wang et al. (2018 & 2019), Mathieson et al. (2018) and Olalde et al. (2018), and others like Lazaridis et al. (2016), Lazaridis et al. (2017), Mittnik et al. (2018), Lamnidis et al. (2018), Fernandes et al. (2018), Jeong et al. (2019), Olalde et al. (2019), etc.
NOTE. As in the Corded Ware ancestry maps, the selected reports in this case are centered on the prototypical Yamnaya ancestry vs. other simplified components, so everything else refers to simplistic ancestral components widespread across populations that do not necessarily share any recent connection, much less a language. In fact, most of the time they clearly didn’t. They can be interpreted as “EHG that is not part of the Yamnaya component”, or “CHG that is not part of the Yamnaya component”. They can’t be read as “expanding EHG people/language” or “expanding CHG people/language”, at least no more than maps of “Steppe ancestry” can be read as “expanding Steppe people/language”. Also, remember that I have left the default behaviour for color classification, so that the highest value (i.e. 1, or white colour) could mean anything from 10% to 100% depending on the specific ancestry and period; that’s what the legend is for… But, fere libenter homines id quod volunt credunt.
Sections:
- Neolithic or the formation of Early Indo-European
- Eneolithic or the expansion of Middle Proto-Indo-European
- Chalcolithic / Early Bronze Age or the expansion of Late Proto-Indo-European
- European Early Bronze Age and MLBA or the expansion of Late PIE dialects
1. Neolithic
Anthony (2019) agrees with the most likely explanation of the CHG component found in Yamnaya, as derived from steppe hunter-fishers close to the lower Volga basin. The ultimate origin of this specific CHG-like component that eventually formed part of the Pre-Yamnaya ancestry is not clear, though:
The hunter-fisher camps that first appeared on the lower Volga around 6200 BC could represent the migration northward of un-admixed CHG hunter-fishers from the steppe parts of the southeastern Caucasus, a speculation that awaits confirmation from aDNA.
The typical EHG component that formed part eventually of Pre-Yamnaya ancestry came from the Middle Volga Basin, most likely close to the Samara region, as shown by the sampled Samara hunter-gatherer (ca. 5600-5500 BC):
After 5000 BC domesticated animals appeared in these same sites in the lower Volga, and in new ones, and in grave sacrifices at Khvalynsk and Ekaterinovka. CHG genes and domesticated animals flowed north up the Volga, and EHG genes flowed south into the North Caucasus steppes, and the two components became admixed.

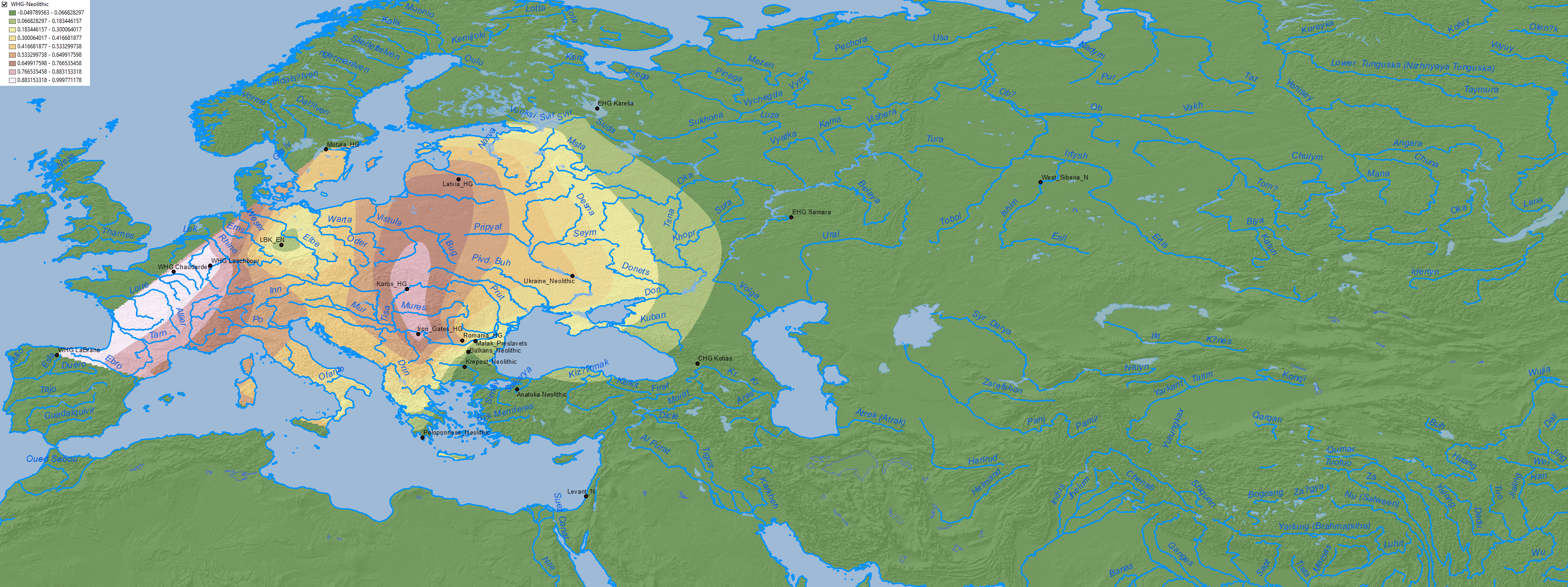
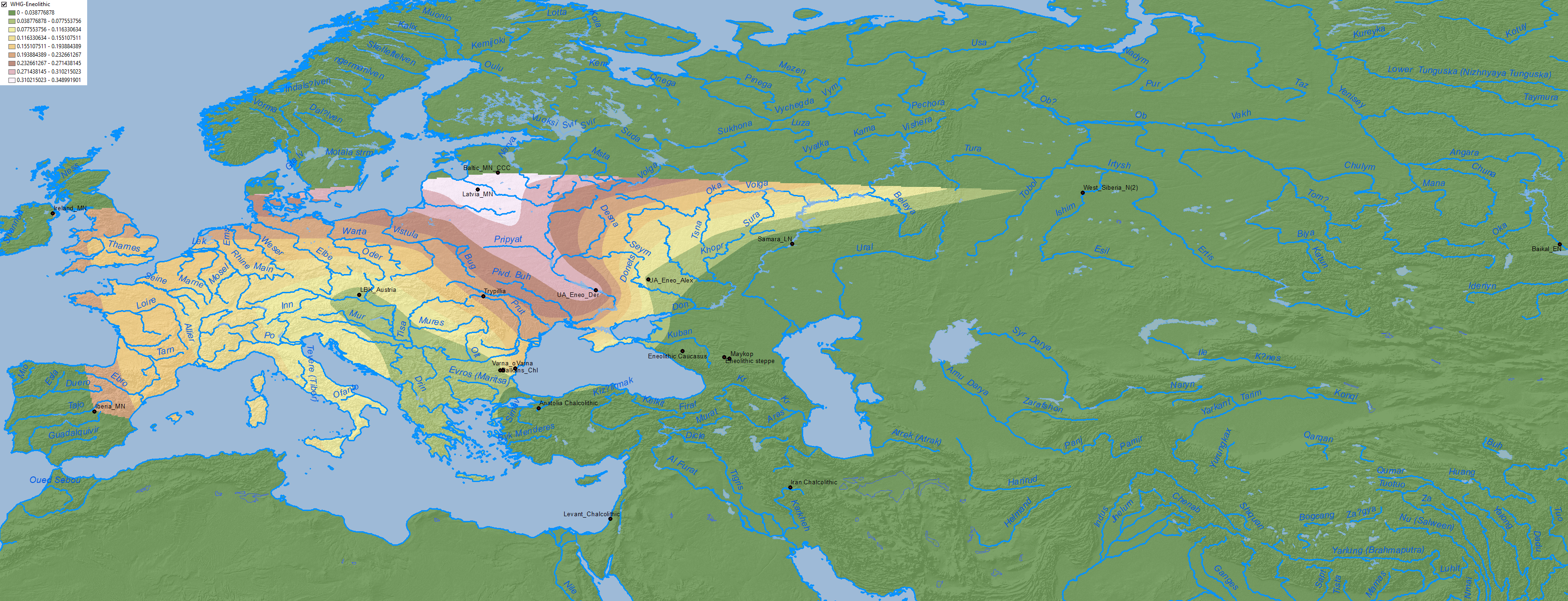
To the west, in the Dnieper-Dniester area, WHG became the dominant ancestry after the Mesolithic, at the expense of EHG, revealing a likely mating network reaching to the north into the Baltic:
Like the Mesolithic and Neolithic populations here, the Eneolithic populations of Dnieper-Donets II type seem to have limited their mating network to the rich, strategic region they occupied, centered on the Rapids. The absence of CHG shows that they did not mate frequently if at all with the people of the Volga steppes (…)
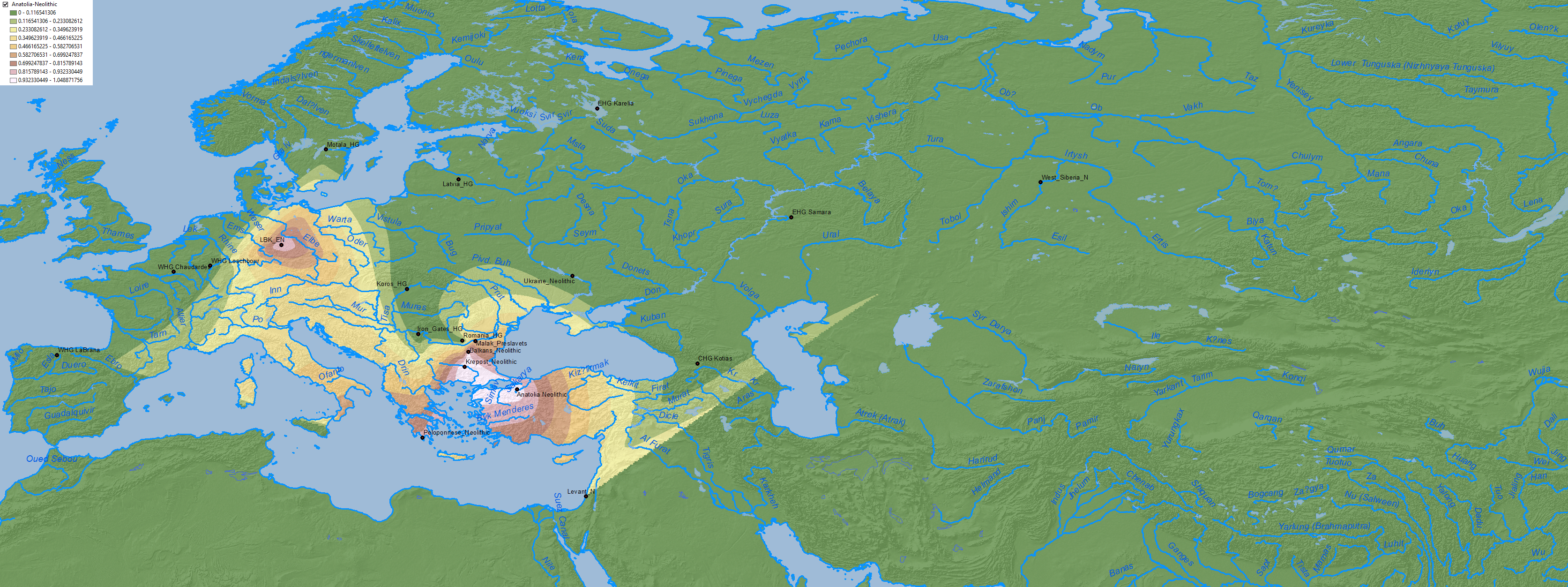
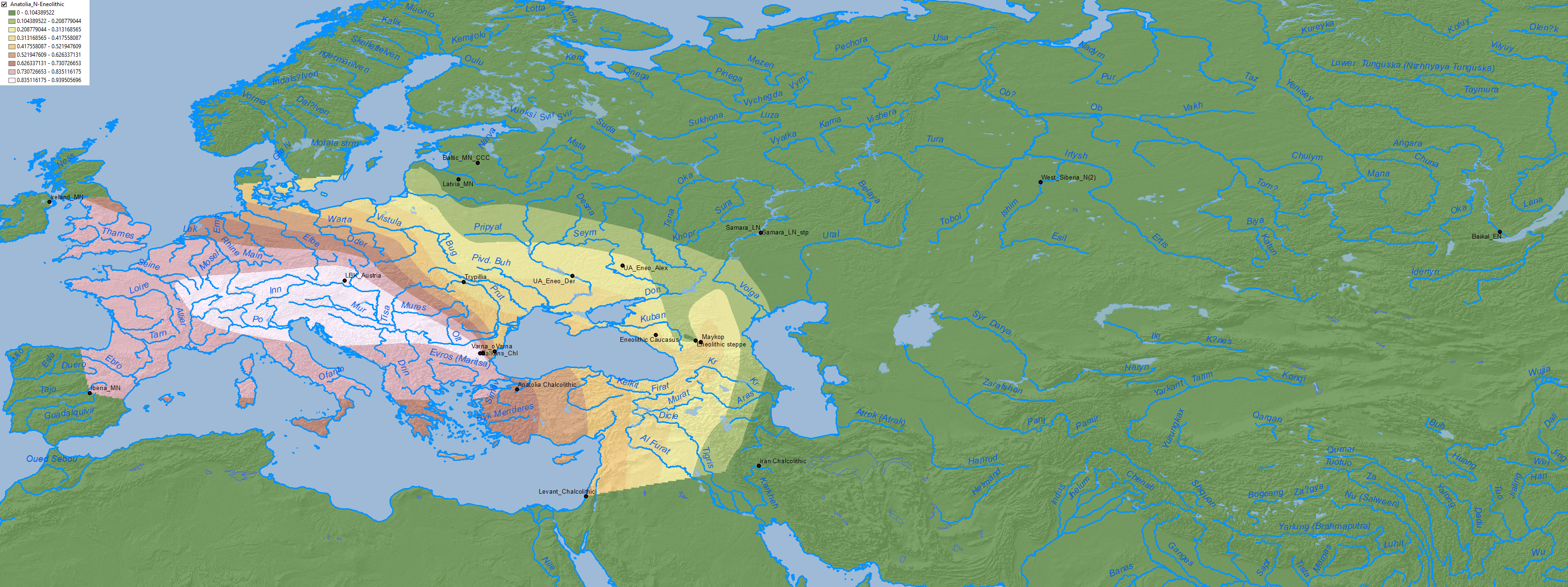
North-West Anatolia Neolithic ancestry, proper of expanding Early European farmers, is found up to border of the Dniester, as Anthony (2007) had predicted.
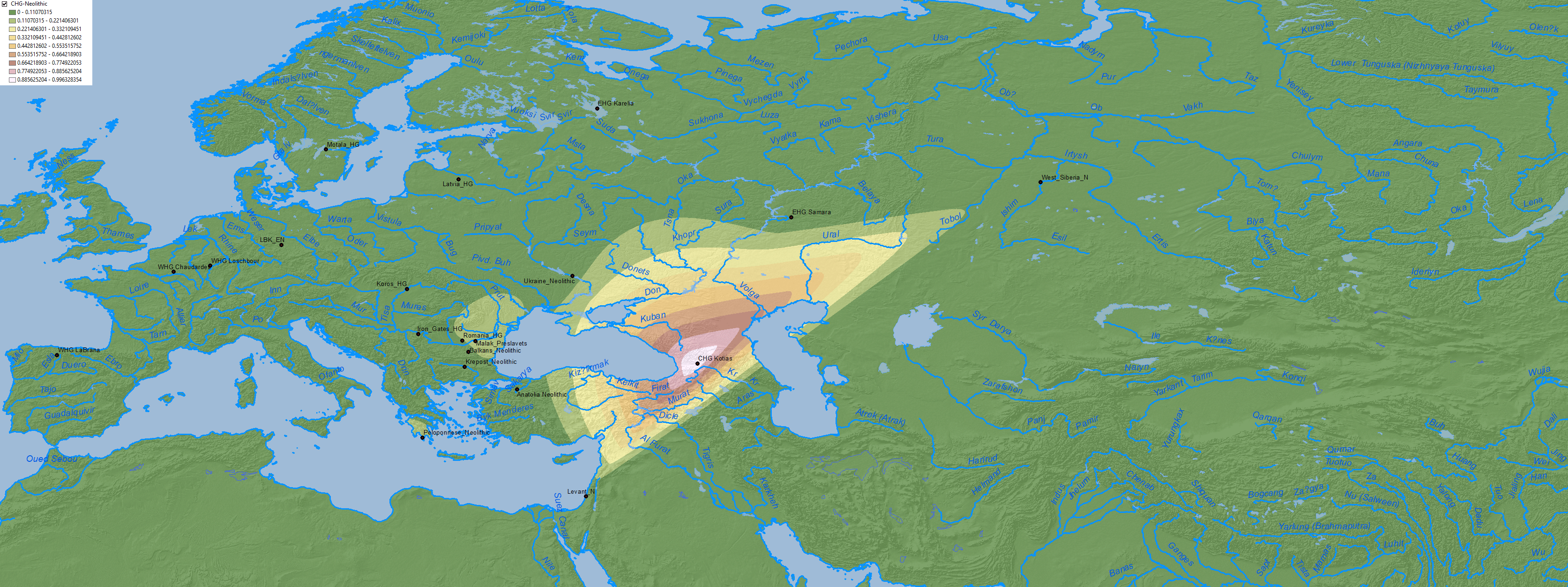
2. Eneolithic
From Anthony (2019):
After approximately 4500 BC the Khvalynsk archaeological culture united the lower and middle Volga archaeological sites into one variable archaeological culture that kept domesticated sheep, goats, and cattle (and possibly horses). In my estimation, Khvalynsk might represent the oldest phase of PIE.
(…) this middle Volga mating network extended down to the North Caucasian steppes, where at cemeteries such as Progress-2 and Vonyuchka, dated 4300 BC, the same Khvalynsk-type ancestry appeared, an admixture of CHG and EHG with no Anatolian Farmer ancestry, with steppe-derived Y-chromosome haplogroup R1b. These three individuals in the North Caucasus steppes had higher proportions of CHG, overlapping Yamnaya. Without any doubt, a CHG population that was not admixed with Anatolian Farmers mated with EHG populations in the Volga steppes and in the North Caucasus steppes before 4500 BC. We can refer to this admixture as pre-Yamnaya, because it makes the best currently known genetic ancestor for EHG/CHG R1b Yamnaya genomes.
From Wang et al (2019):
Three individuals from the sites of Progress 2 and Vonyuchka 1 in the North Caucasus piedmont steppe (‘Eneolithic steppe’), which harbour EHG and CHG related ancestry, are genetically very similar to Eneolithic individuals from Khvalynsk II and the Samara region. This extends the cline of dilution of EHG ancestry via CHG-related ancestry to sites immediately north of the Caucasus foothills
{kind=link}
NOTE. Unpublished samples from Ekaterinovka have been previously reported as within the R1b-L23 tree. Interestingly, although the Varna outlier is a female, the Balkan outlier from Smyadovo shows two positive SNP calls for hg. R1b-M269. However, its poor coverage makes its most conservative haplogroup prediction R-M343.
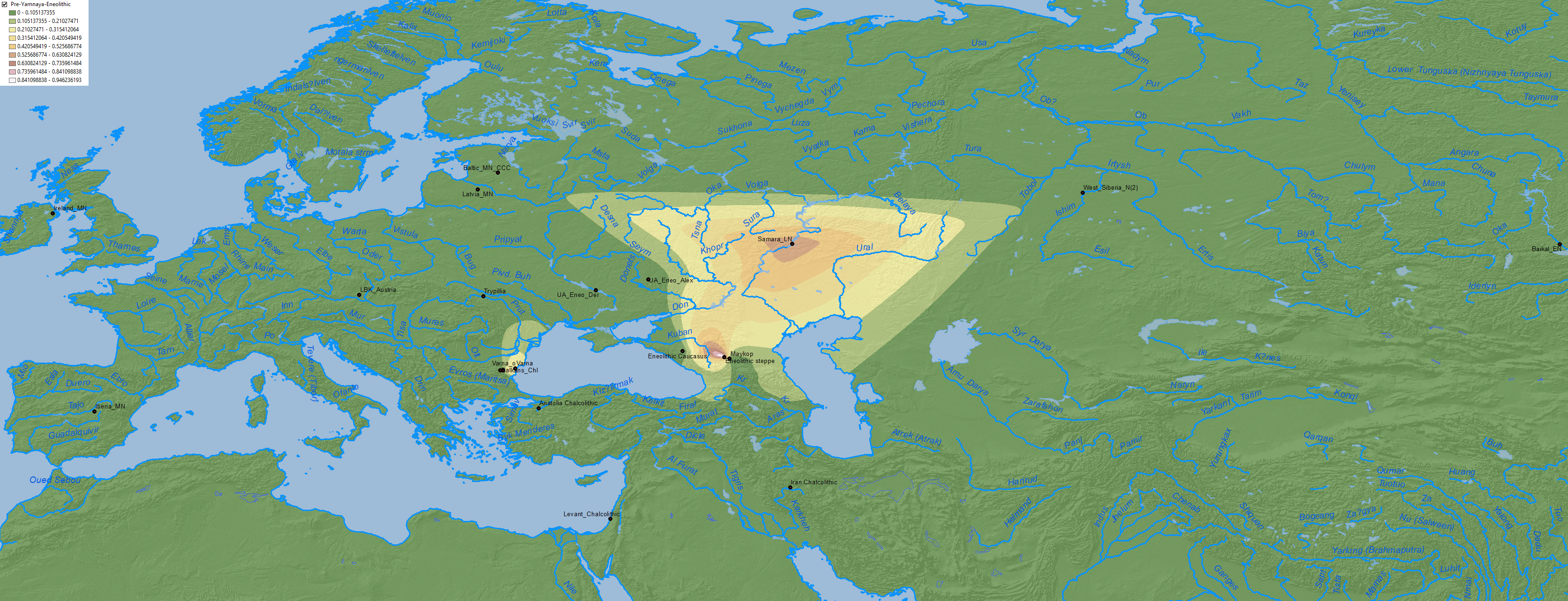
The formation of this Pre-Yamnaya ancestry sets this Volga-Caucasus Khvalynsk community apart from the rest of the EHG-like population of eastern Europe.
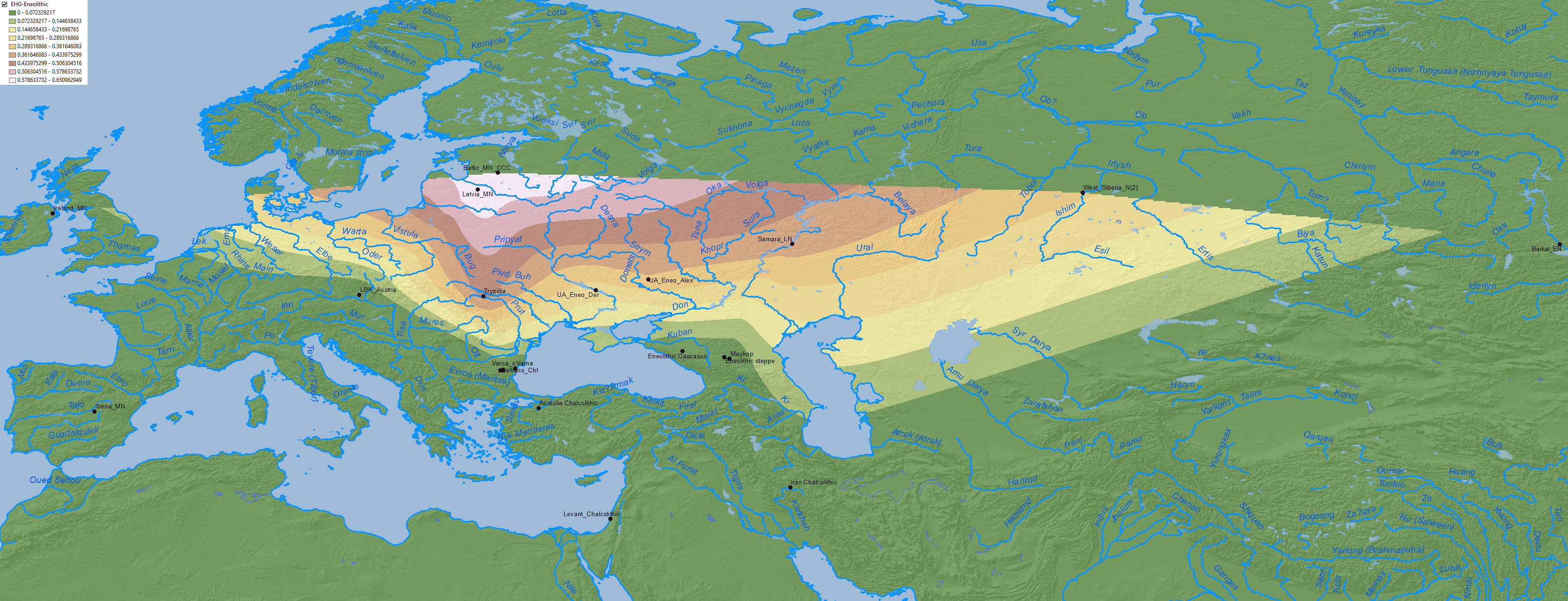
Anthony (2019) seems to rely on ADMIXTURE graphics when he writes that the late Sredni Stog sample from Alexandria shows “80% Khvalynsk-type steppe ancestry (CHG&EHG)”. While this seems the most logical conclusion of what might have happened after the Suvorovo-Novodanilovka expansion through the North Pontic steppes (see my post on “Steppe ancestry” step by step), formal stats have not confirmed that.
In fact, analyses published in Wang et al. (2019) rejected that Corded Ware groups are derived from this Pre-Yamnaya ancestry, a reality that had been already hinted in Narasimhan et al. (2018), when Steppe_EMBA showed a poor fit for expanding Srubna-Andronovo populations. Hence the need to consider the whole CHG component of the North Pontic area separately:
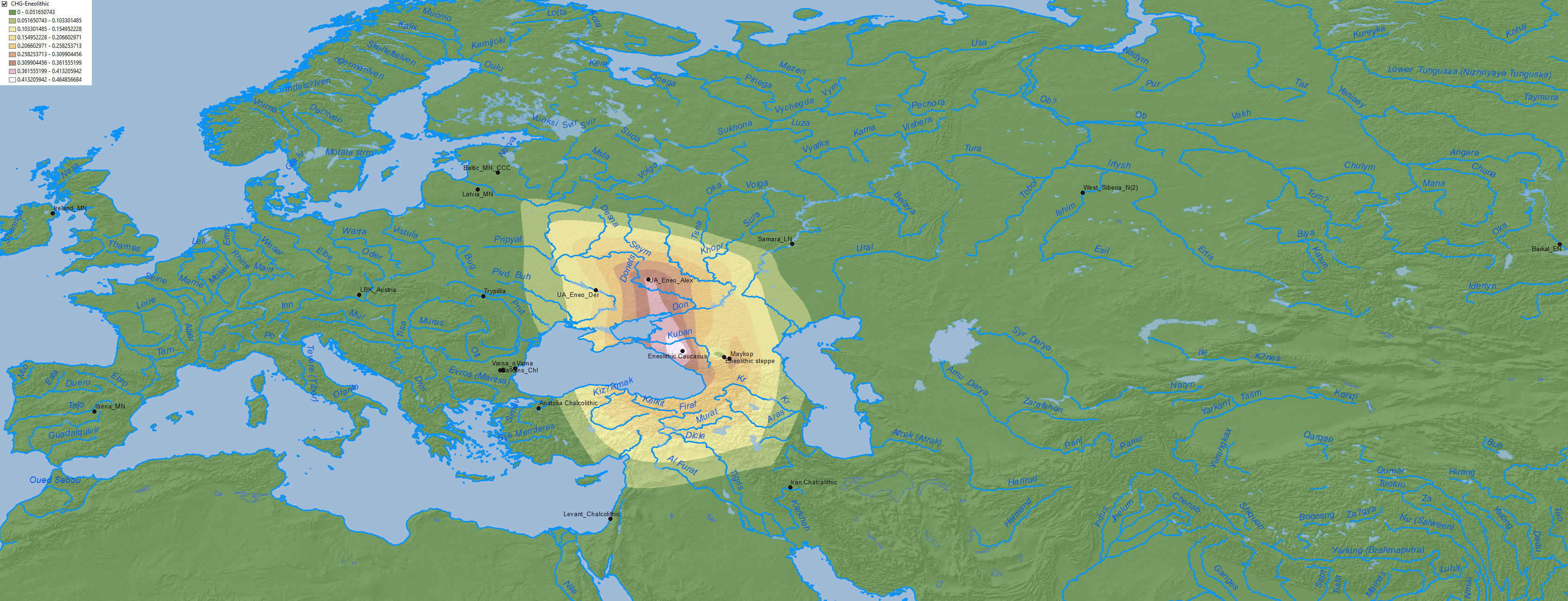
NOTE. Fits for WHG + CHG + EHG in Neolithic and Eneolithic populations are taken in part from Mathieson et al. (2019) supplementary materials (download Excel here). Unfortunately, while data on the Ukraine_Eneolithic outlier from Alexandria abounds, I don’t have specific data on the so-called ‘outlier’ from Dereivka compared to the other two analyzed together, so these maps of CHG and EHG expansion are possibly showing a lesser distribution to the west than the real one ca. 4000-3500 BC.
Anatolia Neolithic ancestry clearly spread to the east into the north Pontic area through a Middle Eneolithic mating network, most likely opened after the Khvalynsk expansion:
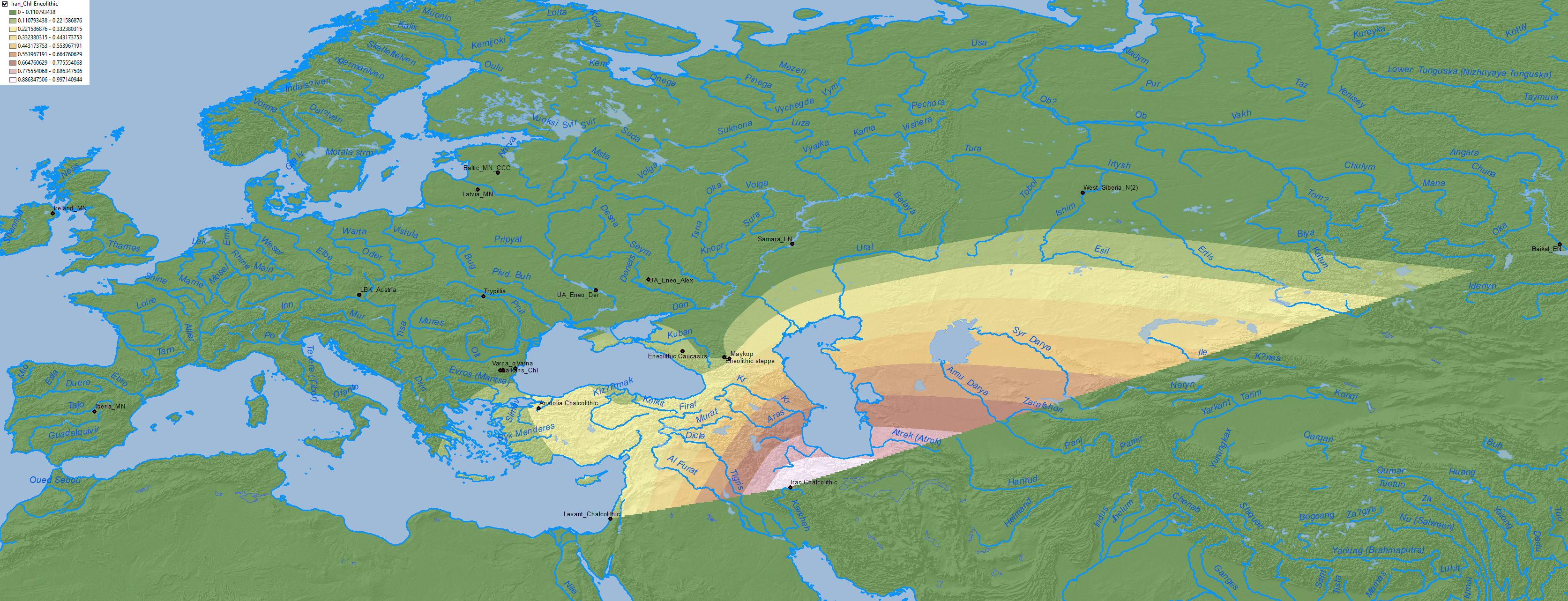
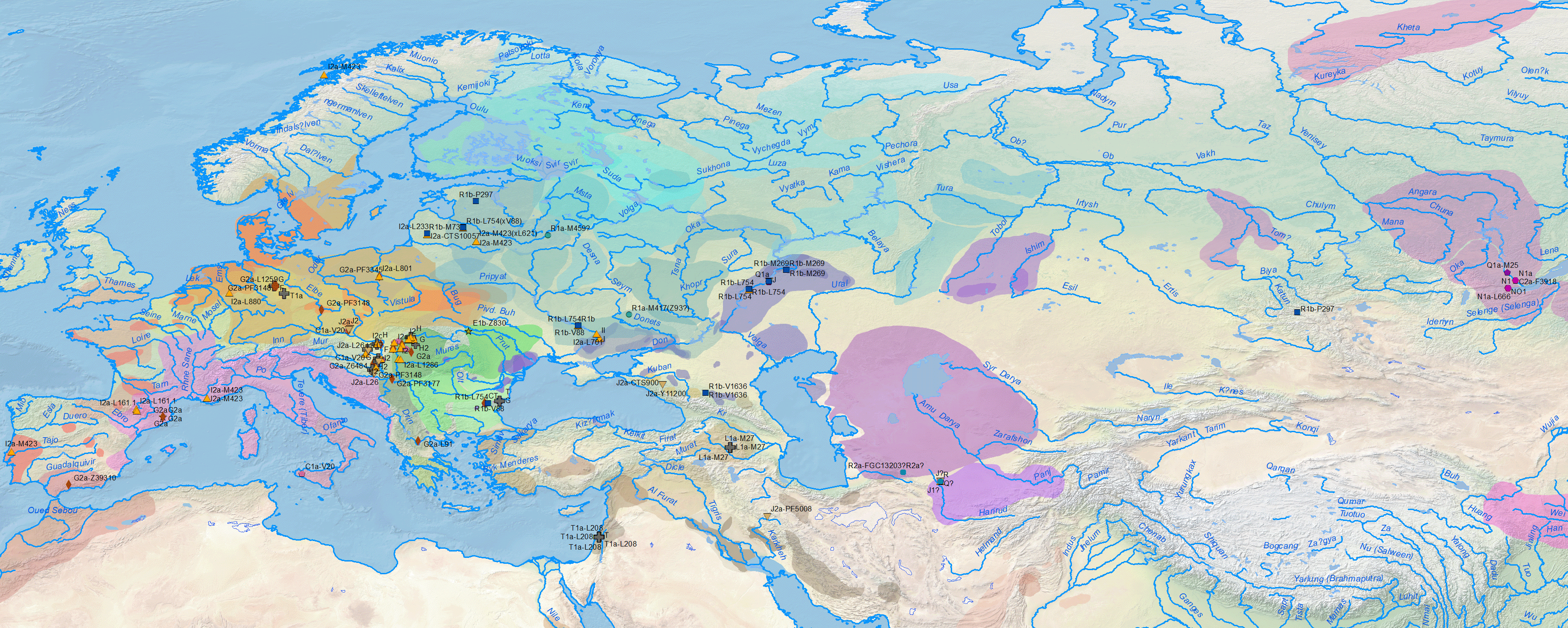
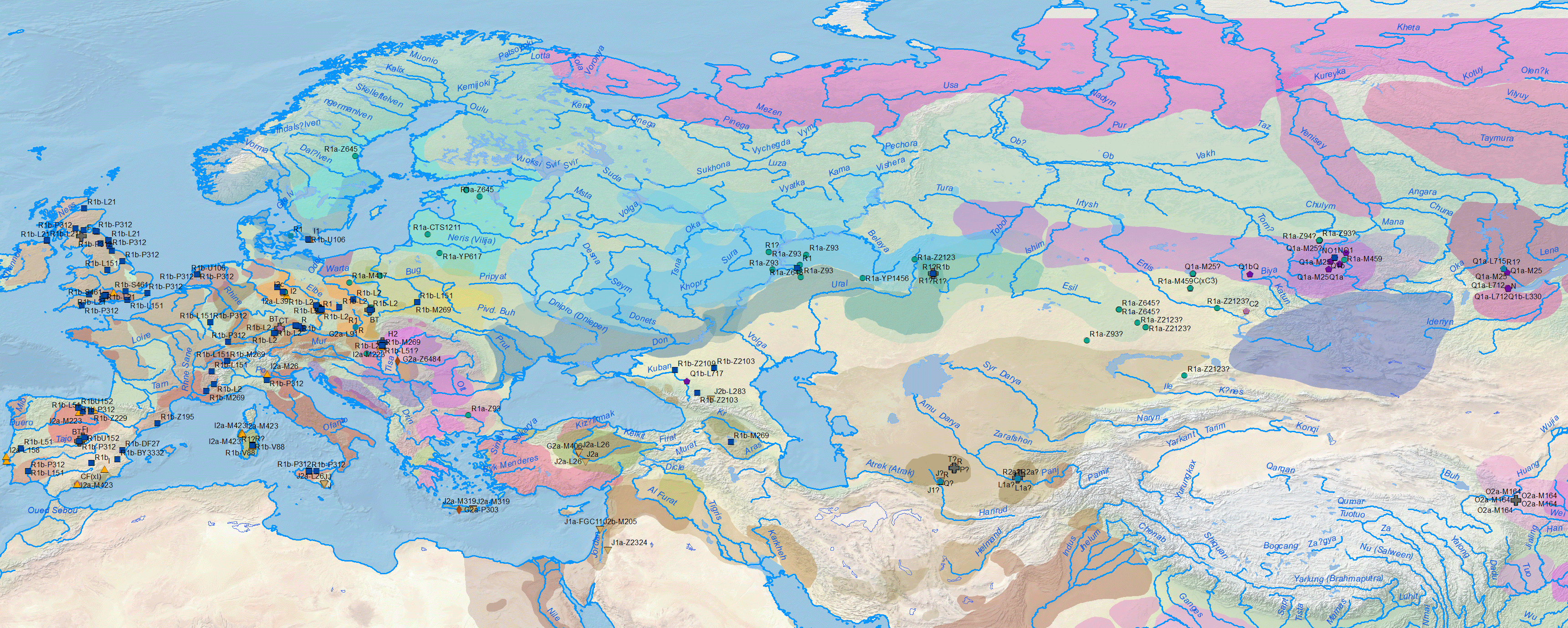
Regarding Y-chromosome haplogroups, Anthony (2019) insists on the evident association of Khvalynsk, Yamnaya, and the spread of Pre-Yamnaya and Yamnaya ancestry with the expansion of elite R1b-L754 (and some I2a2) individuals:
3. Early Bronze Age
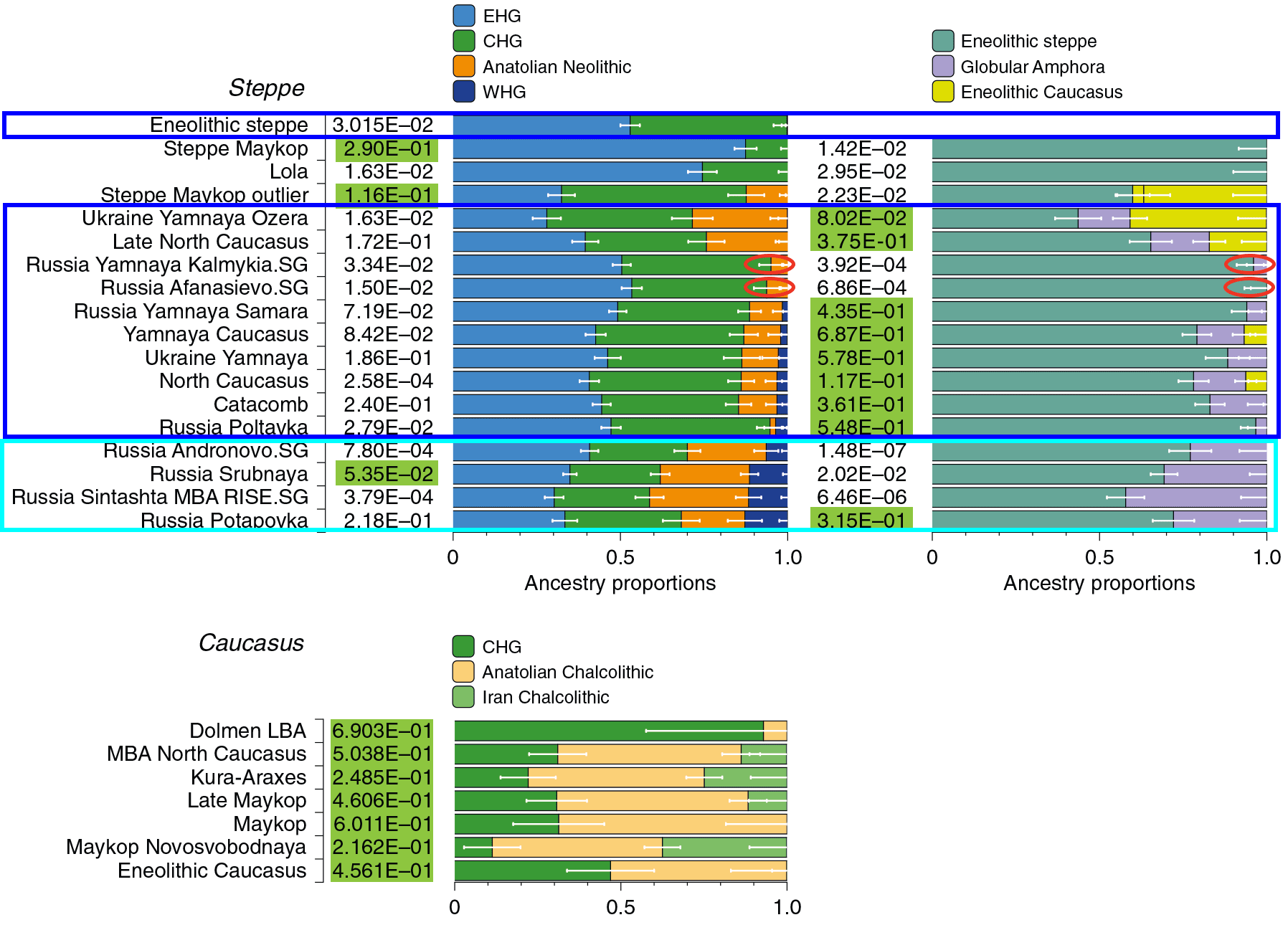
Data from Wang et al. (2019) show that Corded Ware-derived populations do not have good fits for Eneolithic_Steppe-like ancestry, no matter the model. In other words: Corded Ware populations show not only a higher contribution of Anatolia Neolithic ancestry (ca. 20-30% compared to the ca. 2-10% of Yamnaya); they show a different EHG + CHG combination compared to the Pre-Yamnaya one.
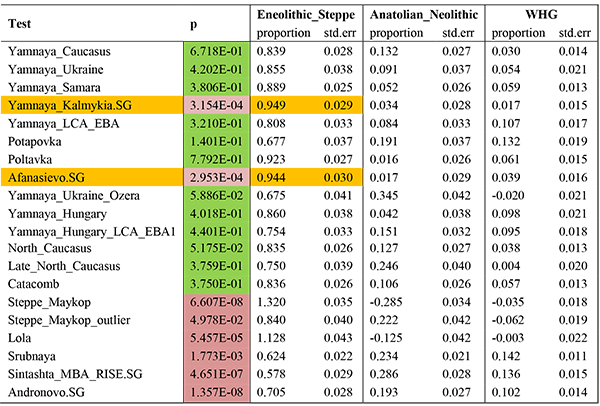
Left populations: Test, Eneolithic_steppe, Anatolian_Neolithic, WHG.
Right populations: Mbuti.DG, Ust_Ishim.DG, Kostenki14, MA1, Han.DG, Papuan.DG, Onge.DG, Villabruna, Vestonice16, ElMiron, Ethiopia_4500BP.SG, Karitiana.DG, Natufian, Iran_Ganj_Dareh_Neolithic.
Yamnaya Kalmykia and Afanasievo show the closest fits to the Eneolithic population of the North Caucasian steppes, rejecting thus sizeable contributions from Anatolia Neolithic and/or WHG, as shown by the SD values. Both probably show then a Pre-Yamnaya ancestry closest to the late Repin population.
EBA maps include data from Wang et al. (2018) supplementary materials, specifically unpublished Yamnaya samples from Hungary that appeared in analysis of the preprint, but which were taken out of the definitive paper. Their location among Yamnaya settlers from Hungary is speculative, although most uncovered kurgans in Hungary are concentrated in the Tisza-Danube interfluve.
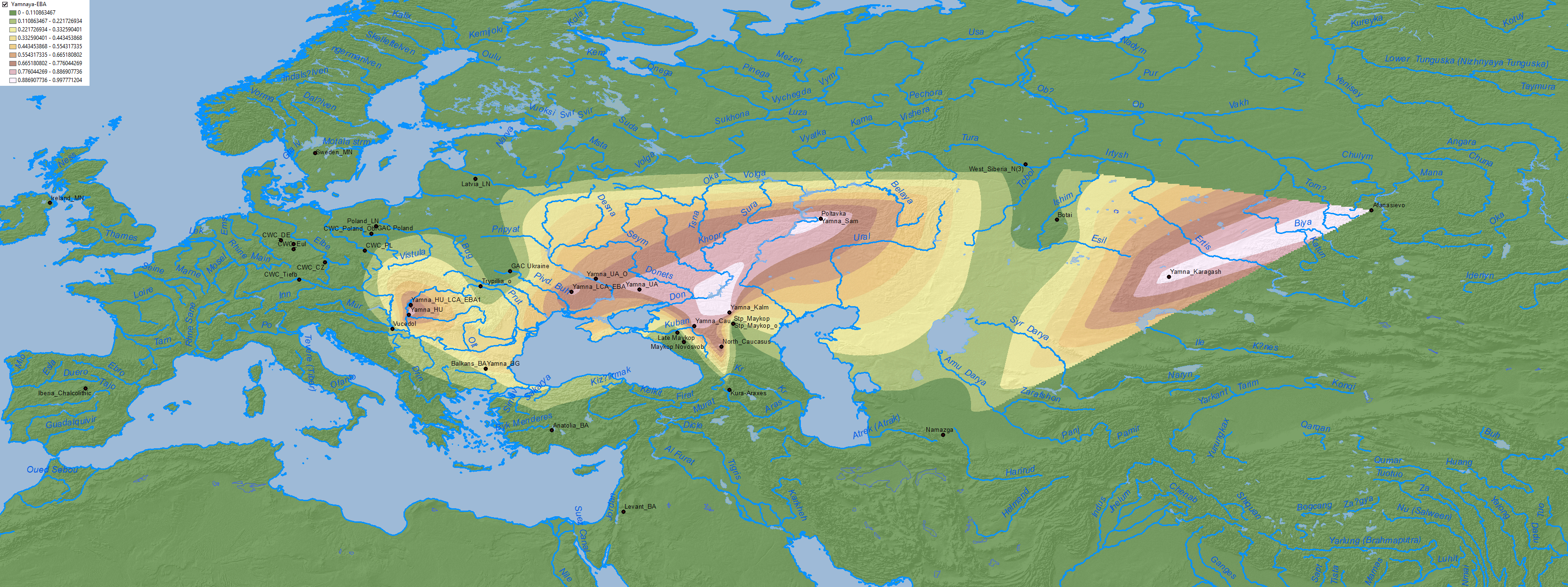
The Y-chromosome bottleneck of elite males from Proto-Indo-European clans under R1b-L754 and some I2a2 subclades, already visible in the Khvalynsk sampling, became even more noticeable in the subsequent expansion of late Repin/early Yamnaya elites under R1b-L23 and I2a-L699:

Maps of CHG, EHG, Anatolia Neolithic, and probably WHG show the expansion of these components among Corded Ware-related groups in North Eurasia, apart from other cultures close to the Caucasus:
NOTE. For maps with actual formal stats of Corded Ware ancestry from the Early Bronze Age to the modern times, you can read the post Corded Ware ancestry in North Eurasia and the Uralic expansion.
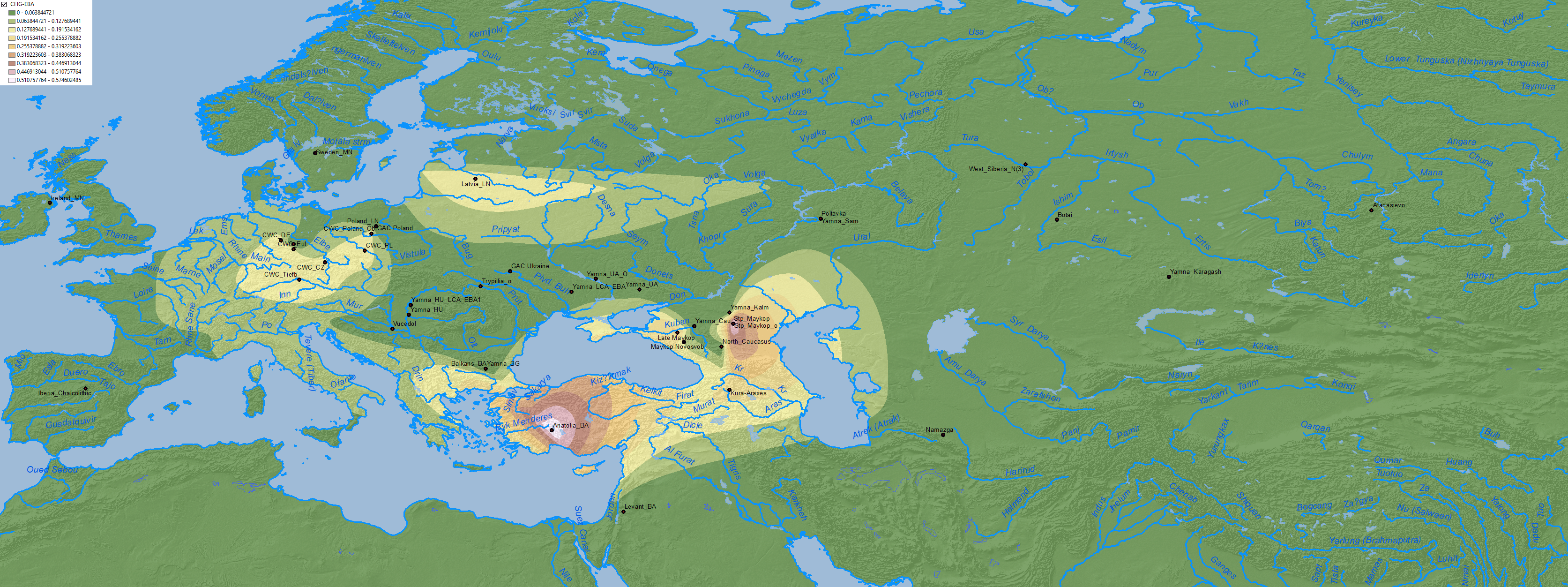
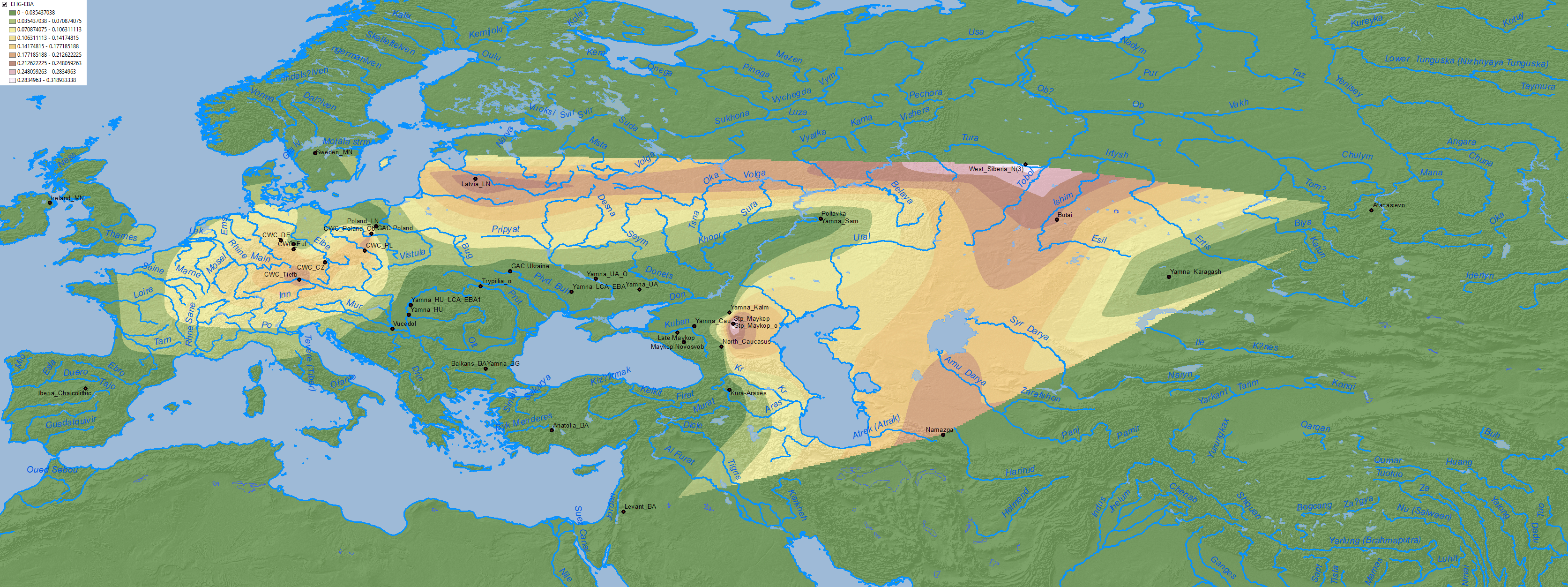
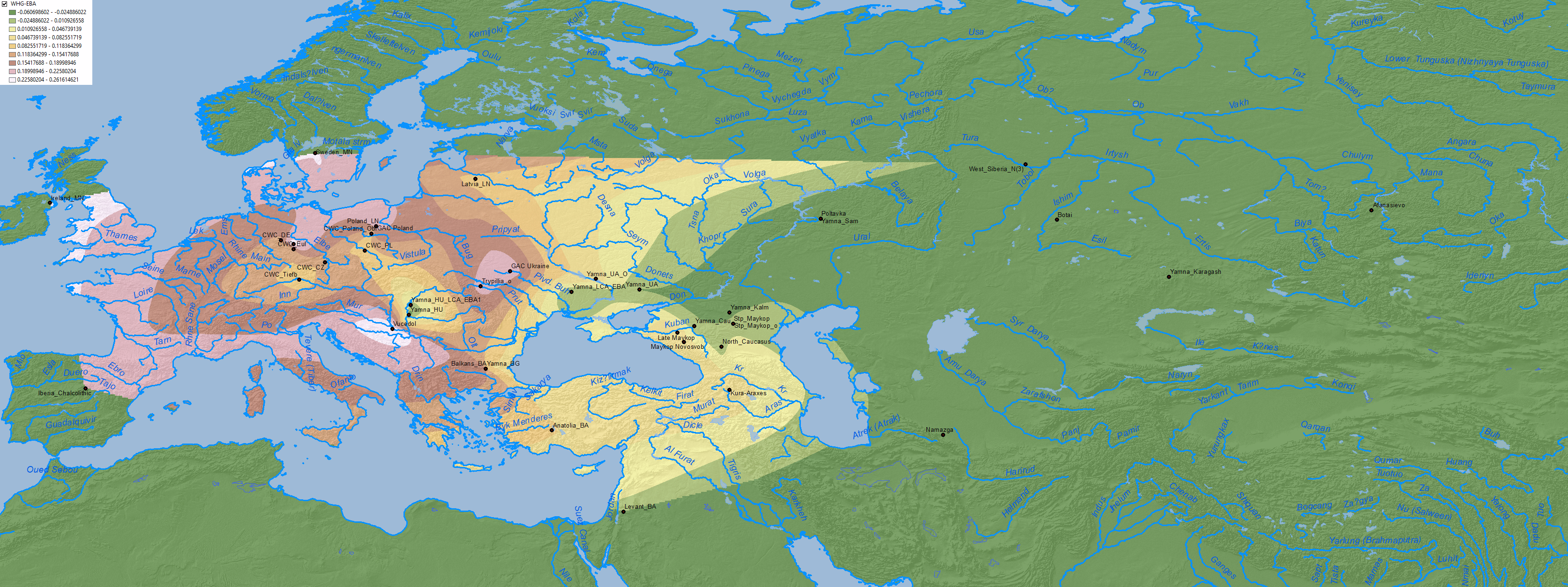

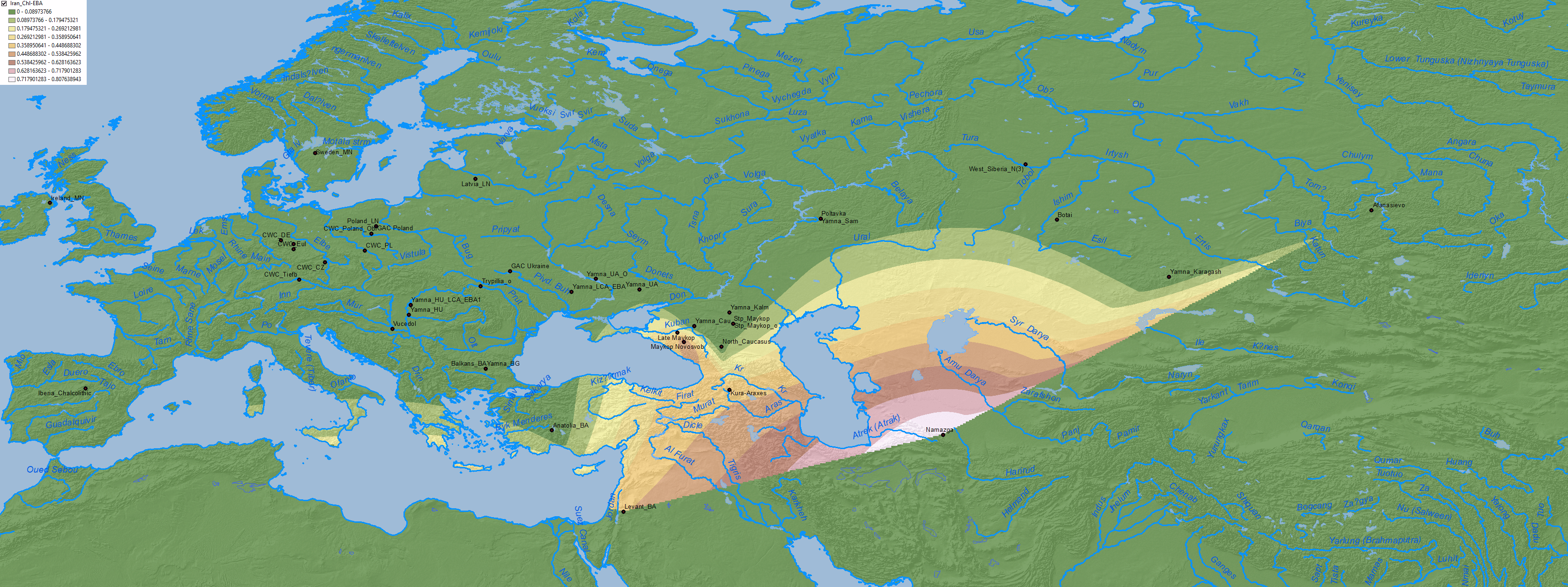
4. Middle to Late Bronze Age
The following maps show the most likely distribution of Yamnaya ancestry during the Bell Beaker-, Balkan-, and Sintashta-Potapovka-related expansions.
4.1. Bell Beakers
The amount of Yamnaya ancestry is probably overestimated among populations where Bell Beakers replaced Corded Ware. A map of Yamnaya ancestry among Bell Beakers gets trickier for the following reasons:
- Expanding Repin peoples of Pre-Yamnaya ancestry must have had admixture through exogamy with late Sredni Stog/Proto-Corded Ware peoples during their expansion into the North Pontic area, and Sredni Stog in turn had probably some Pre-Yamnaya admixture, too (although they don’t appear in the simplistic formal stats above). This is supported by the increase of Anatolia farmer ancestry in more western Yamna samples.
- Later, Yamnaya admixed through exogamy with Corded Ware-like populations in Central Europe during their expansion. Even samples from the Middle to Upper Danube and around the Lower Rhine will probably show increasing contributions of Steppe_MLBA, at the same time as they show an increasing proportion of EEF-related ancestry.
- To complicate things further, the late Corded Ware Espersted family (from ca. 2500 BC or later) shows, in turn, what seems like a recent admixture with Yamnaya vanguard groups, with the sample of highest Yamnaya ancestry being the paternal uncle of other individuals (all of hg. R1a-M417), suggesting that there might have been many similar Central European mating networks from the mid-3rd millennium BC on, of (mainly) Yamnaya-like R1b elites displaying a small proportion of CW-like ancestry admixing through exogamy with Corded Ware-like peoples who already had some Yamnaya ancestry.
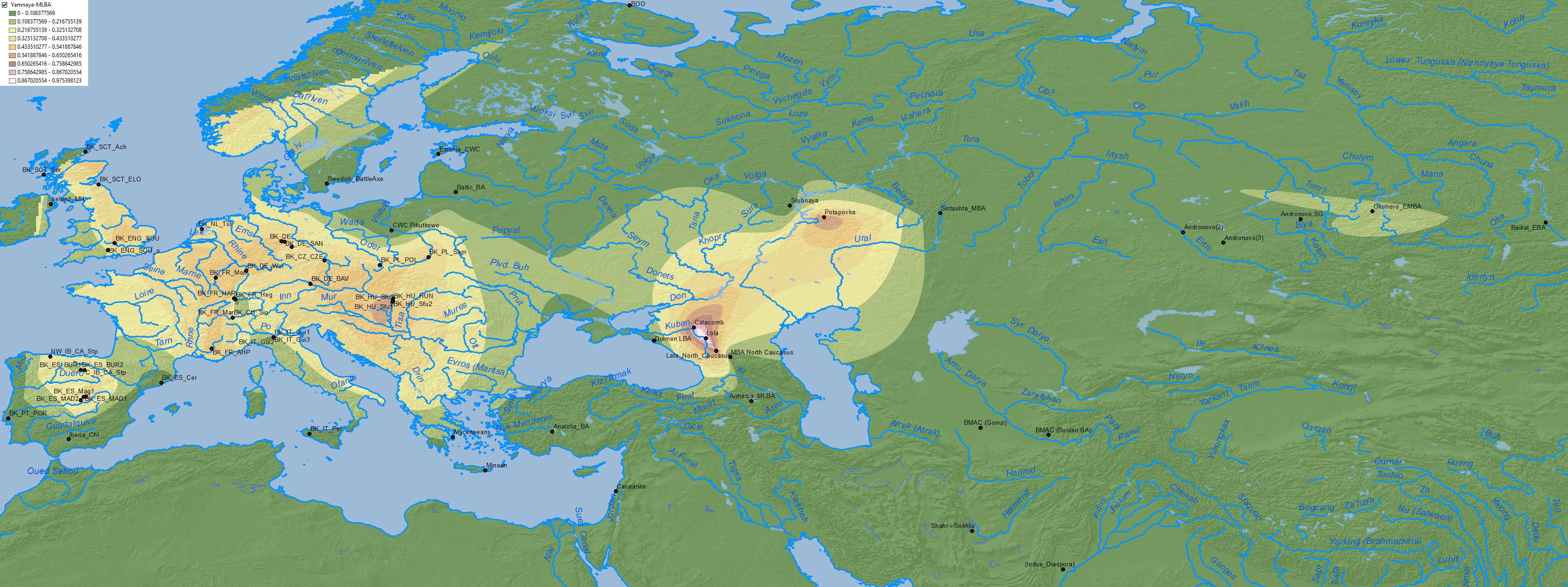
NOTE. Terms like “exogamy”, “male-driven migration”, and “sex bias”, are not only based on the Y-chromosome bottlenecks visible in the different cultural expansions since the Palaeolithic. Despite the scarce sampling available in 2017 for analysis of “Steppe ancestry”-related populations, it appeared to show already a male sex bias in Goldberg et al. (2017), and it has been confirmed for Neolithic and Copper Age population movements in Mathieson et al. (2018) – see Supplementary Table 5. The analysis of male-biased expansion of “Steppe ancestry” in CWC Esperstedt and Bell Beaker Germany is, for the reasons stated above, not very useful to distinguish their mutual influence, though.
Based on data from Olalde et al. (2019), Bell Beakers from Germany are the closest sampled ones to expanding East Bell Beakers, and those close to the Rhine – i.e. French, Dutch, and British Beakers in particular – show a clear excess “Steppe ancestry” due to their exogamy with local Corded Ware groups:
Only one 2-way model fits the ancestry in Iberia_CA_Stp with P-value>0.05: Germany_Beaker + Iberia_CA. Finding a Bell Beaker-related group as a plausible source for the introduction of steppe ancestry into Iberia is consistent with the fact that some of the individuals in the Iberia_CA_Stp group were excavated in Bell Beaker associated contexts. Models with Iberia_CA and other Bell Beaker groups such as France_Beaker (P-value=7.31E-06), Netherlands_Beaker (P-value=1.03E-03) and England_Beaker (P-value=4.86E-02) failed, probably because they have slightly higher proportions of steppe ancestry than the true source population.

The exogamy with Corded Ware-like groups in the Lower Rhine Basin seems at this point undeniable, as is the origin of Bell Beakers around the Middle-Upper Danube Basin from Yamnaya Hungary.
To avoid this excess “Steppe ancestry” showing up in the maps, since Bell Beakers from Germany pack the most Yamnaya ancestry among East Bell Beakers outside Hungary (ca. 51.1% “Steppe ancestry”), I equated this maximum with BK_Scotland_Ach (which shows ca. 61.1% “Steppe ancestry”, highest among western Beakers), and applied a simple rule of three for “Steppe ancestry” in Dutch and British Beakers.
NOTE. Formal stats for “Steppe ancestry” in Bell Beaker groups are available in Olalde et al. (2018) supplementary materials (PDF). I didn’t apply this adjustment to Bk_FR groups because of the R1b Bell Beaker sample from the Champagne/Alsace region reported by Samantha Brunel that will pack more Yamnaya ancestry than any other sampled Beaker to date, hence probably driving the Yamnaya ancestry up in French samples.
The most likely outcome in the following years, when Yamnaya and Corded Ware ancestry are investigated separately, is that Yamnaya ancestry will be much lower the farther away from the Middle and Lower Danube region, similar to the case in Iberia, so the map above probably overestimates this component in most Beakers to the north of the Danube. Even the late Hungarian Beaker samples, who pack the highest Yamnaya ancestry (up to 75%) among Beakers, represent likely a back-migration of Moravian Beakers, and will probably show a contribution of Corded Ware ancestry due to the exogamy with local Moravian groups.
Despite this decreasing admixture as Bell Beakers spread westward, the explosive expansion of Yamnaya R1b male lineages (in words of David Reich) and the radical replacement of local ones – whether derived from Corded Ware or Neolithic groups – shows the true extent of the North-West Indo-European expansion in Europe:

4.2. Palaeo-Balkan
There is scarce data on Palaeo-Balkan movements yet, although it is known that:
- Yamnaya ancestry appears among Mycenaeans, with the Yamnaya Bulgaria sample being its best current ancestral fit;
- the emergence of steppe ancestry and R1b-M269 in the eastern Mediterranean was associated with Ancient Greeks;
- Thracians, Albanians, and Armenians also show R1b-M269 subclades and “Steppe ancestry”.
4.3. Sintashta-Potapovka-Filatovka
Interestingly, Potapovka is the only Corded Ware derived culture that shows good fits for Yamnaya ancestry, despite having replaced Poltavka in the region under the same Corded Ware-like (Abashevo) influence as Sintashta.
This proves that there was a period of admixture in the Pre-Proto-Indo-Iranian community between CWC-like Abashevo and Yamnaya-like Catacomb-Poltavka herders in the Sintashta-Potapovka-Filatovka community, probably more easily detectable in this group because of the specific temporal and geographic sampling available.
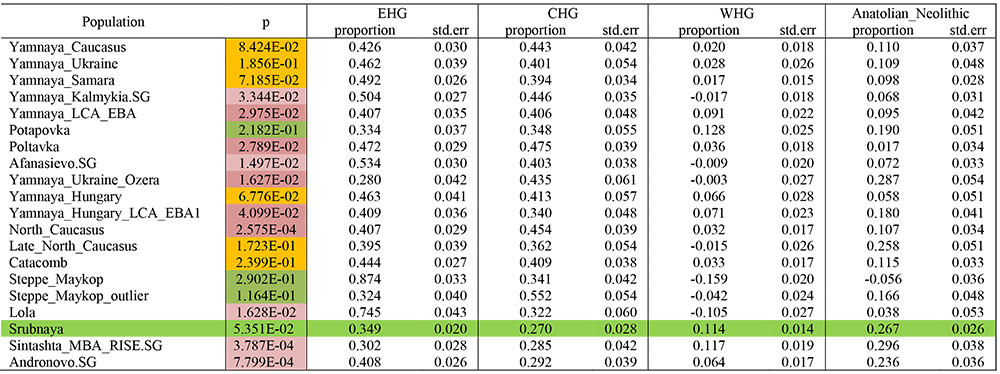
Left populations: Steppe cluster, EHG, CHG, WHG, Anatolian_Neolithic
Right populations: Mbuti.DG, Ust_Ishim.DG, Kostenki14, MA1, Han.DG, Papuan.DG, Onge.DG, Villabruna, Vestonice16, ElMiron, Ethiopia_4500BP.SG, Karitiana.DG, Natufian, Iran_Ganj_Dareh_Neolithic.
Srubnaya ancestry shows a best fit with non-Pre-Yamnaya ancestry, i.e. with different CHG + EHG components – possibly because the more western Potapovka (ancestral to Proto-Srubnaya Pokrovka) also showed good fits for it. Srubnaya shows poor fits for Pre-Yamnaya ancestry probably because Corded Ware-like (Abashevo) genetic influence increased during its formation.
On the other hand, more eastern Corded Ware-derived groups like Sintashta and its more direct offshoot Andronovo show poor fits with this model, too, but their fits are still better than those including Pre-Yamnaya ancestry.
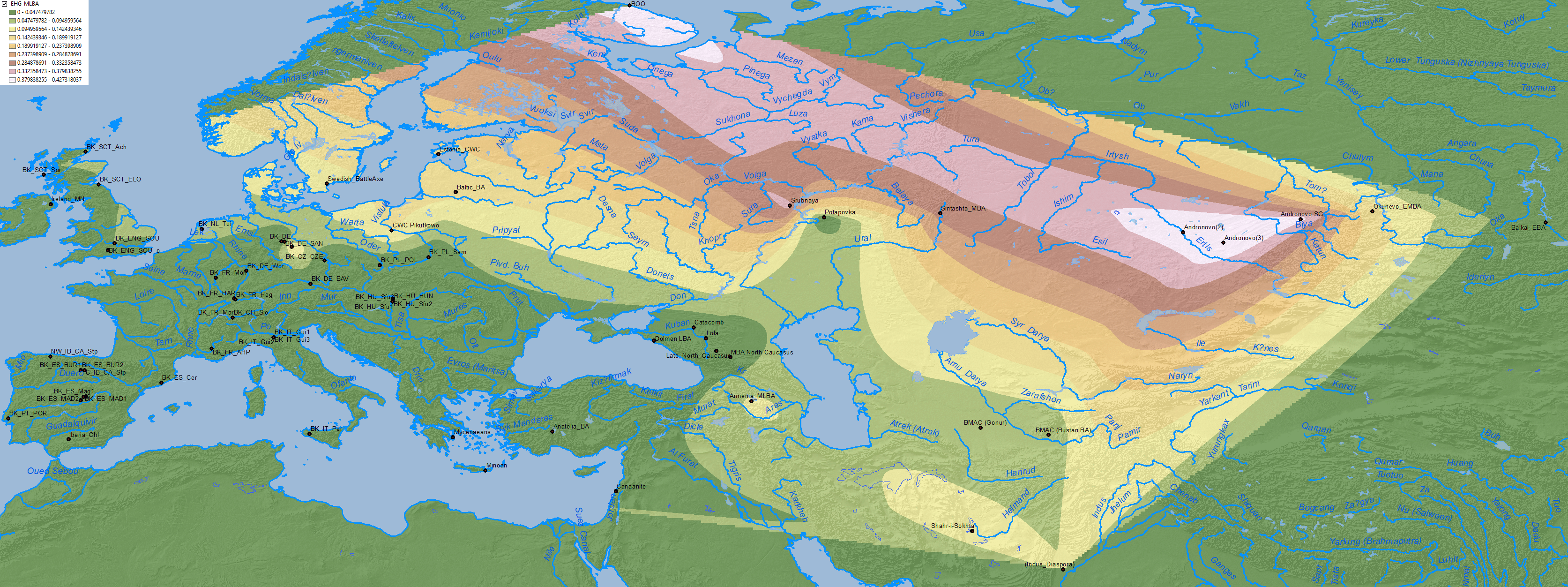
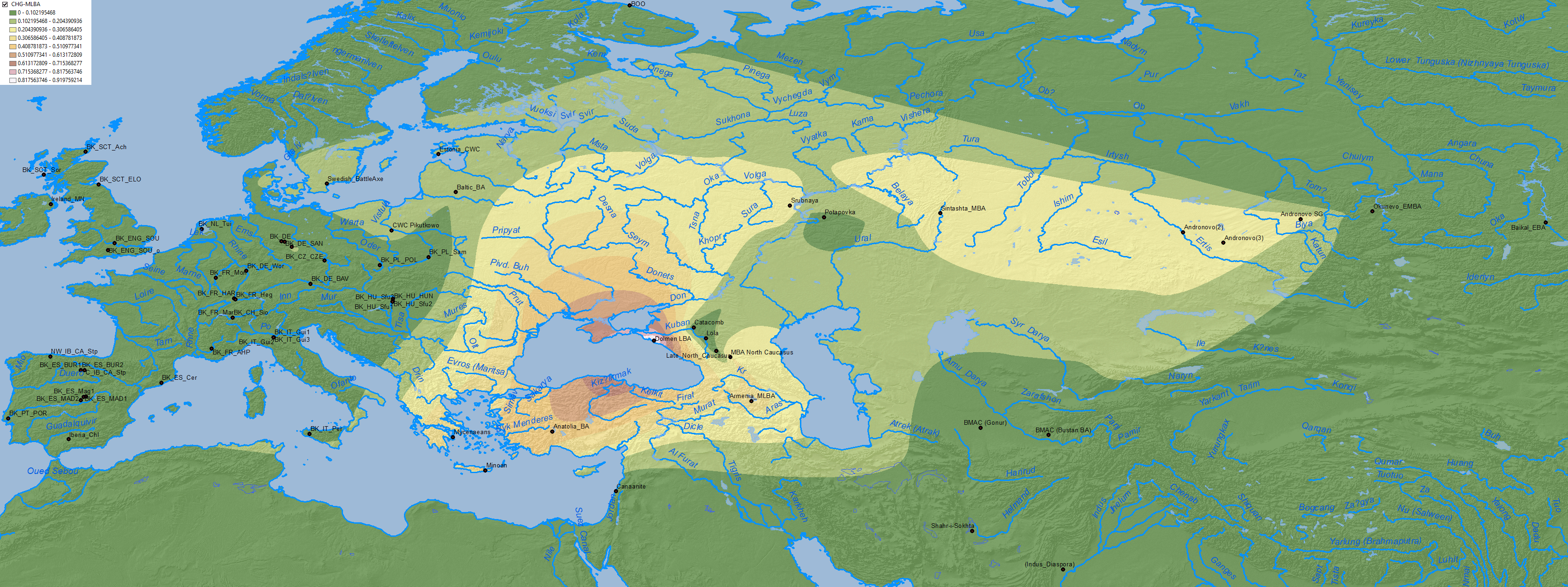

NOTE For maps with actual formal stats of Corded Ware ancestry from the Early Bronze Age to the modern times, you should read the post Corded Ware ancestry in North Eurasia and the Uralic expansion instead.
The bottleneck of Proto-Indo-Iranians under R1a-Z93 was not yet complete by the time when the Sintashta-Potapovka-Filatovka community expanded with the Srubna-Andronovo horizon:
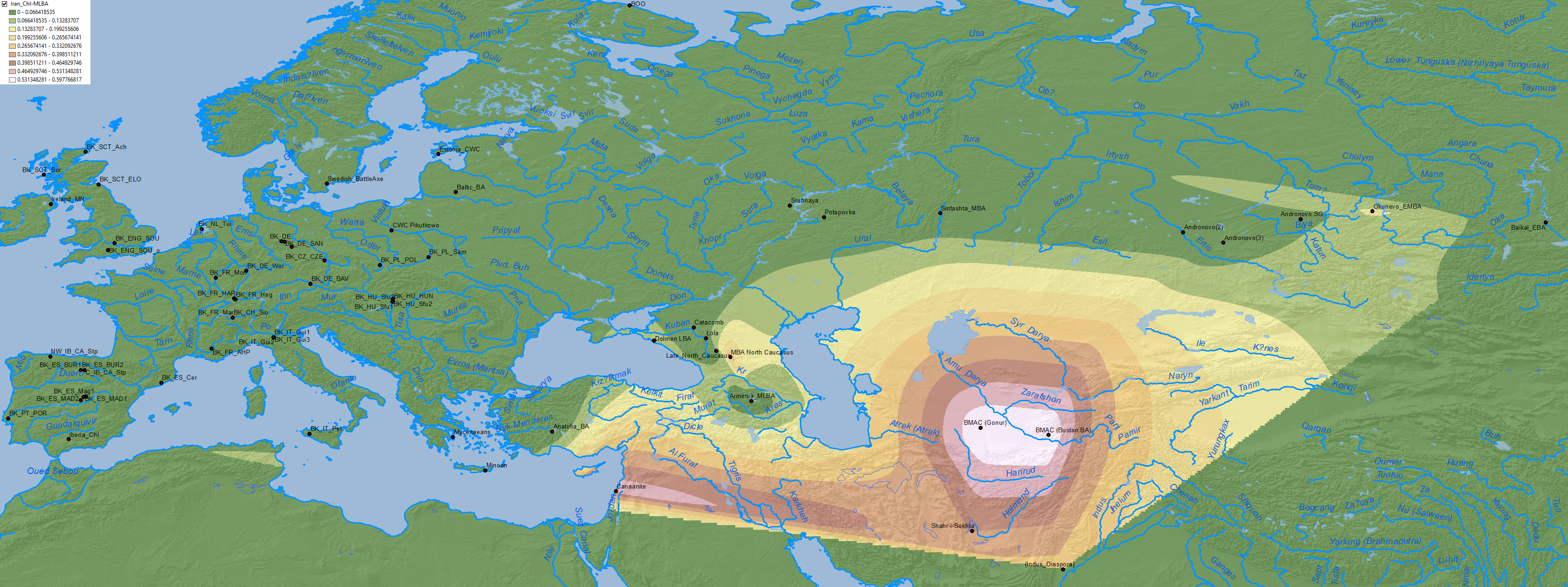
4.4. Afanasevo
At the end of the Afanasevo culture, at least three samples show hg. Q1b (ca. 2900-2500 BC), which seemed to point to a resurgence of local lineages, despite continuity of the prototypical Pre-Yamnaya ancestry. On the other hand, Anthony (2019) makes this cryptic statement:
Yamnaya men were almost exclusively R1b, and pre-Yamnaya Eneolithic Volga-Caspian-Caucasus steppe men were principally R1b, with a significant Q1a minority.
Since the only available samples from the Khvalynsk community are R1b (x3), Q1a(x1), and R1a(x1), it seems strange that Anthony would talk about a “significant minority”, unless Q1a (potentially Q1b in the newer nomenclature) will pop up in some more individuals of those ca. 30 new to be published. Because he also mentions I2a2 as appearing in one elite burial, it seems Q1a (like R1a-M459) will not appear under elite kurgans, although it is still possible that hg. Q1a was involved in the expansion of Afanasevo to the east.
Okunevo, which replaced Afanasevo in the Altai region, shows a majority of hg. Q1b, but also some R1b-M269 samples proper of Afanasevo, suggesting partial genetic continuity.
NOTE. Other sampled Siberian populations clearly show a variety of Q subclades that likely expanded during the Palaeolithic, such as Baikal EBA samples from Ust’Ida and Shamanka with a majority of Q1b, and hg. Q reported from Elunino, Sagsai, Khövsgöl, and also among peoples of the Srubna-Andronovo horizon (the Krasnoyarsk MLBA outlier), and in Karasuk.
From Damgaard et al. Science (2018):
(…) in contrast to the lack of identifiable admixture from Yamnaya and Afanasievo in the CentralSteppe_EMBA, there is an admixture signal of 10 to 20% Yamnaya and Afanasievo in the Okunevo_EMBA samples, consistent with evidence of western steppe influence. This signal is not seen on the X chromosome (qpAdm P value for admixture on X 0.33 compared to 0.02 for autosomes), suggesting a male-derived admixture, also consistent with the fact that 1 of 10 Okunevo_EMBA males carries a R1b1a2a2 Y chromosome related to those found in western pastoralists. In contrast, there is no evidence of western steppe admixture among the more eastern Baikal region region Bronze Age (~2200 to 1800 BCE) samples.
This Yamnaya ancestry has been also recently found to be the best fit for the Iron Age population of Shirenzigou in Xinjiang – where Tocharian languages were attested centuries later – despite the haplogroup diversity acquired during their evolution, likely through an intermediate Chemurchek culture (see a recent discussion on the elusive Proto-Tocharians).
Haplogroup diversity seems to be common in Iron Age populations all over Eurasia, most likely due to the spread of different types of sociopolitical structures where alliances played a more relevant role in the expansion of peoples. A well-known example of this is the spread of Akozino warrior-traders in the whole Baltic region under a partial N1a-VL29-bottleneck associated with the emerging chiefdom-based systems under the influence of expanding steppe nomads.

Surprisingly, then, Proto-Tocharians from Shirenzigou pack up to 74% Yamnaya ancestry, in spite of the 2,000 years that separate them from the demise of the Afanasevo culture. They show more Yamnaya ancestry than any other population by that time, being thus a sort of Late PIE fossils not only in their archaic dialect, but also in their genetic profile:
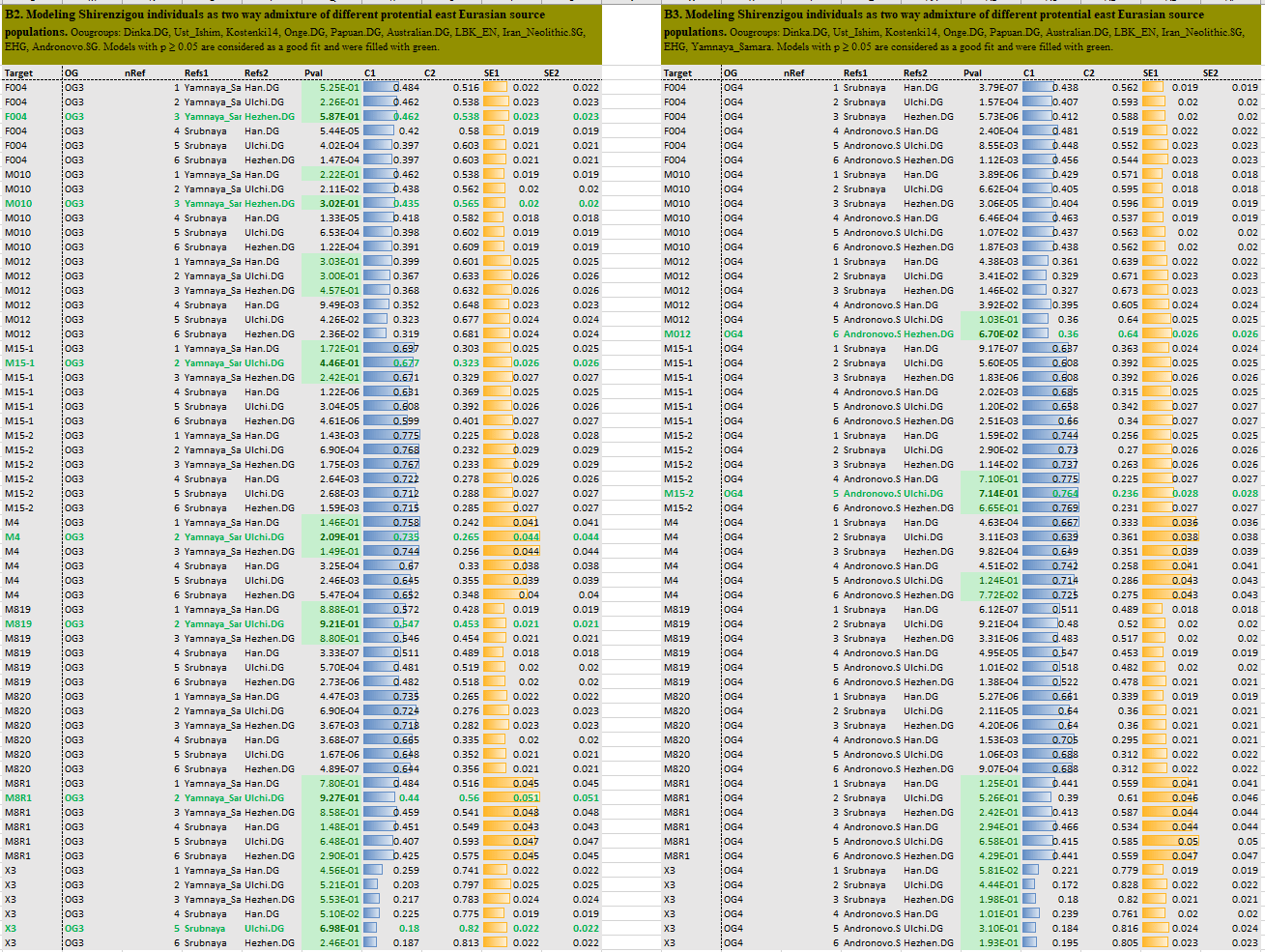
The recent intrusion of Corded Ware-like ancestry, as well as the variable admixture with Siberian and East Asian populations, both point to the known intense Old Iranian and Old/Middle Chinese contacts. The scarce Proto-Samoyedic and Proto-Turkic loans in Tocharian suggest a rather loose, probably more distant connection with East Uralic and Altaic peoples from the forest-steppe and steppe areas to the north (read more about external influences on Tocharian).
Interestingly, both R1b samples, MO12 and M15-2 – likely of Asian R1b-PH155 branch – show a best fit for Andronovo/Srubna + Hezhen/Ulchi ancestry, suggesting a likely connection with Iranians to the east of Xinjiang, who later expanded as the Wusun and Kangju. How they might have been related to Huns and Xiongnu individuals, who also show this haplogroup, is yet unknown, although Huns also show hg. R1a-Z93 (probably most R1a-Z2124) and Steppe_MLBA ancestry, earlier associated with expanding Iranian peoples of the Srubna-Andronovo horizon.
All in all, it seems that prehistoric movements explained through the lens of genetic research fit perfectly well the linguistic reconstruction of Proto-Indo-European and Proto-Uralic.
Related
- Volga Basin R1b-rich Proto-Indo-Europeans of (Pre-)Yamnaya ancestry
- Corded Ware ancestry in North Eurasia and the Uralic expansion
- “Steppe ancestry” step by step: Khvalynsk, Sredni Stog, Repin, Yamna, Corded Ware
- The genetic and cultural barrier of the Pontic-Caspian steppe – forest-steppe ecotone
- Steppe and Caucasus Eneolithic: the new keystones of the EHG-CHG-ANE ancestry in steppe groups
- Kurgan origins and expansion with Khvalynsk-Novodanilovka chieftains
- On the origin of haplogroup R1b-L51 in late Repin / early Yamna settlers
- Bronze Age cultures in the Tarim Basin and the elusive Proto-Tocharians
- Sintashta diet and economy based on domesticated animal products and wild resources
- About Scepters, Horses, and War: on Khvalynsk migrants in the Caucasus and the Danube
- A study of genetic diversity of three isolated populations in Xinjiang using Y-SNP
- Aquitanians and Iberians of haplogroup R1b are exactly like Indo-Iranians and Balto-Slavs of haplogroup R1a
- Consequences of Damgaard et al. 2018 (II): The late Khvalynsk migration waves with R1b-L23 lineages
- The origin of social complexity in the development of the Sintashta culture
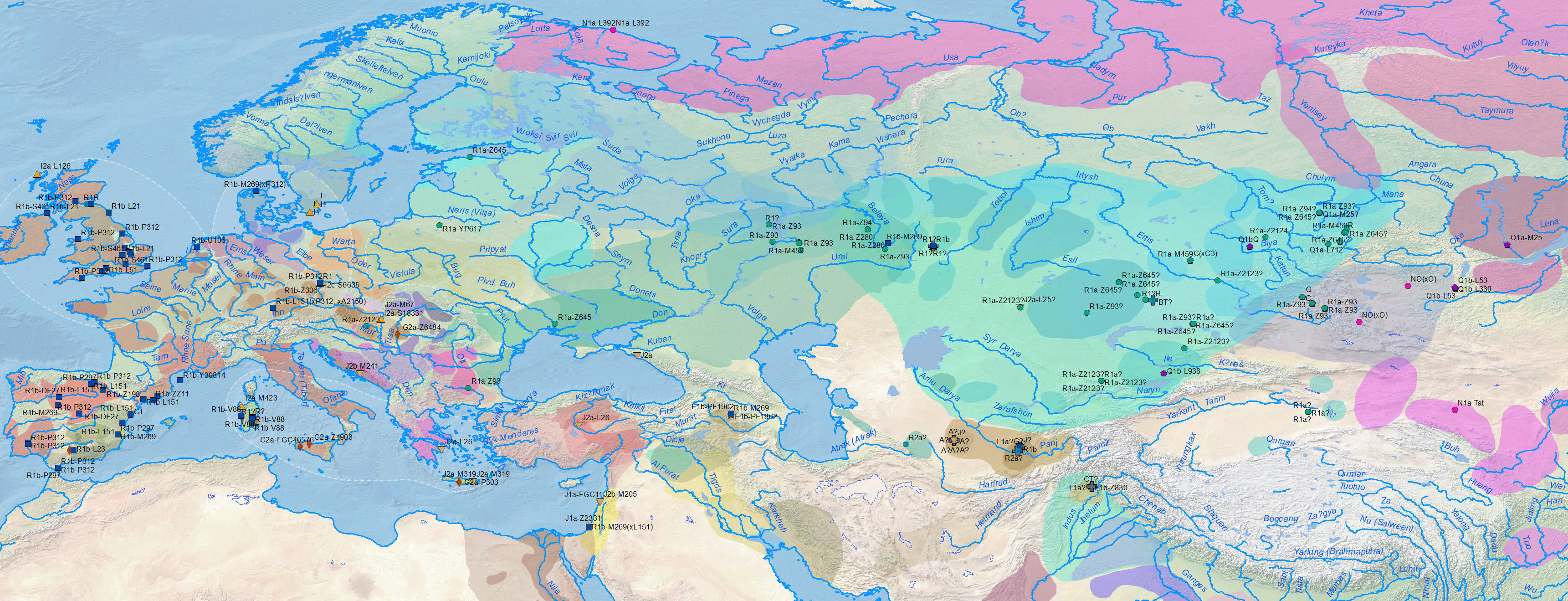
- On the origin and spread of haplogroup R1a-Z645 from eastern Europe
- North Pontic steppe Eneolithic cultures, and an alternative Indo-Slavonic model