New paper (behind paywall) Paternal origin of Paleo-Indians in Siberia: insights from Y-chromosome sequences by Wei et al., Eur. J. Hum. Genet. (2018)
Interesting excerpts (for Eurasian migrations):
Differentiation and diffusion in Palaeolithic Siberia
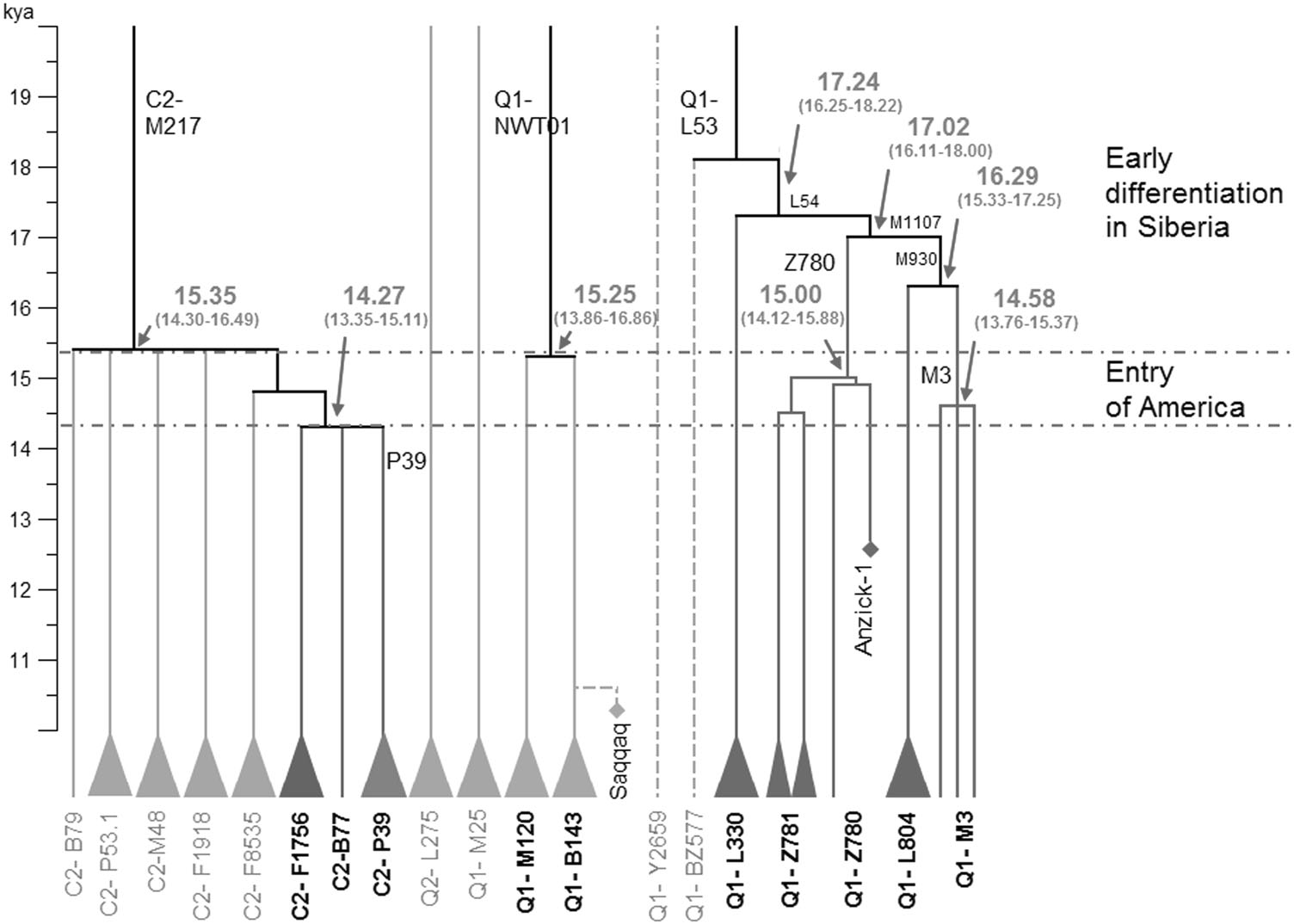
Based on the phylogenetic analyses and the current distributions of relative sub-lineages, we propose that the prehistoric population differentiation in Siberia after the LGM (post-LGM) provided the genetic basis for the emergence of the Paleo-Indian, American aborigine, population. According to the phylogenetic tree of Y-chromosome haplogroup C2-M217 (Fig. 2 and Figure S1), eight sub-lineages emerged in a short period between 15.3 kya and 14.3 kya (Table S5). Within these sub-lineages, haplogroups C2-M48, C2-F1918, and C2- F1756 are predominant paternal lineages in modern Altaic-speaking populations [46, 51, 52]. Samples of haplogroups C2-F8535 and C2-P53.1 were found in two Turkic- and Mongolic-speaking minorities in China (Table S1). Both archeological and genetic data suggest that Altaic-speaking populations are results of population expansion in the past several thousand years in the Altai Mountain, Mongolia Plateau, and Amur River region [51–54].
By contrast, three other sub-lineages, C2-B79, C2-B77, and C2-P39, appear only in Koryaks and Native Americans [16, 35]. The latitude of the Altai Mountain, the Mongolia Plateau, and Amur River region are much lower than that of Beringia, where the ancestors of Native Americans finally separated from their close relatives in Siberia. Therefore, the phylogeographic patterns of sub-lineages of C2-M217 in this study reveal a major splitting event between populations in a lower latitude region of Siberia and ancestors of Koryaks and Native Americans during the post-LGM period.
The sub-lineages of the Y-chromosome Q-M242 haplogroup were found in populations throughout the Eurasia continent. According to available data, the Q1-L804 lineage is exclusively found in Northwest Europe, while Q1-M120 is primarily restricted to East Asia [48]. Additionally, the lineage Q1-L330 is the predominant paternal lineage in Altai, Tuva, and Kets in South Siberia [34–36, 55]. A number of Q1-M242 samples have also been found in ancient remains from South Siberia and adjacent regions [56, 57]. Other sub-lineages of Q-M242 are scattered widely in different geographic regions of Eurasia, including Q1-L275, Q1-M25, and Q1-Y2659 [14, 35, 37, 58]. Additionally, the Y-chromosome of a 6000–5100 BCE sample (I4550) from Zvejnieki, Latvia has been identified as Q1-L56 [59]. These findings suggest that the sub-lineages of Q-M242 started to diffuse throughout Eurasia in a very ancient period.
Emergence of Paleo-Indian populations
The revised phylogenetic tree of Y-chromosome haplogroup Q-M242 in this study provides clues regarding the origin of Native American lineages Q1-M3 and Q1-Z780 (Fig. 3). According to our estimates, haplogroup Q1-L54 expanded rapidly between 17.2 kya and 15.0 kya and finally gave rise to two major founding paternal lineages of Native American populations, known as Q1-Z780 and Q1-M3. Ancient DNA studies indicate that the early population in South Siberia, represented by MA1 genomes, had a genetic influence on both modern western European and Native American populations [7]. Therefore, we conclude that the accumulated diversity of sub-lineages of Q-M242 before 15.3 kya resulted from the in situ differentiation of Q-M242 in Central Eurasia and South Siberia since the Paleolithic Age, and the appearance of the Paleo-Indian population is part of the great human diffusion throughout the Eurasia after the Last Glacial Maximum.
The Southern Caucasus PIE homeland
{kind=link}
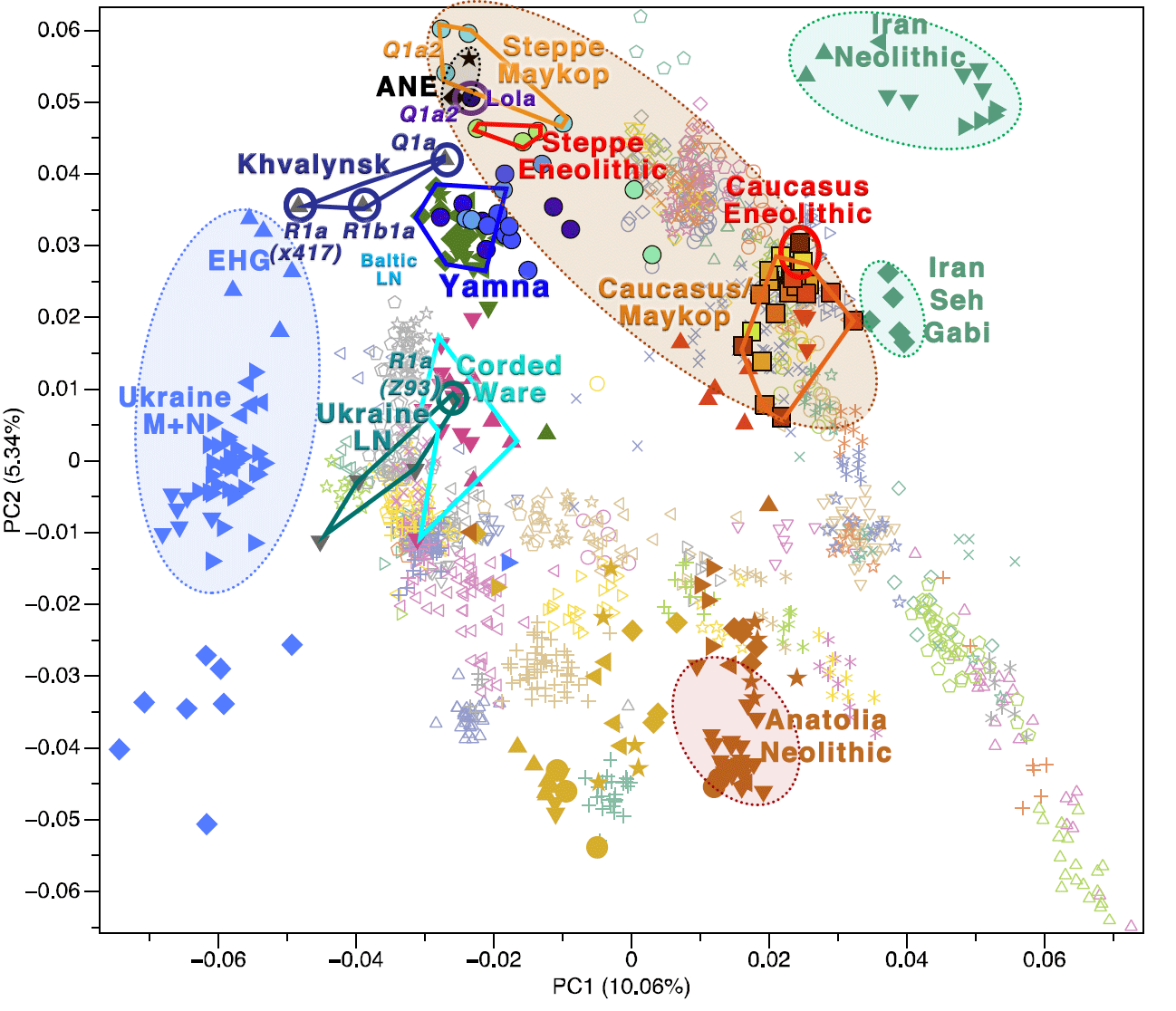
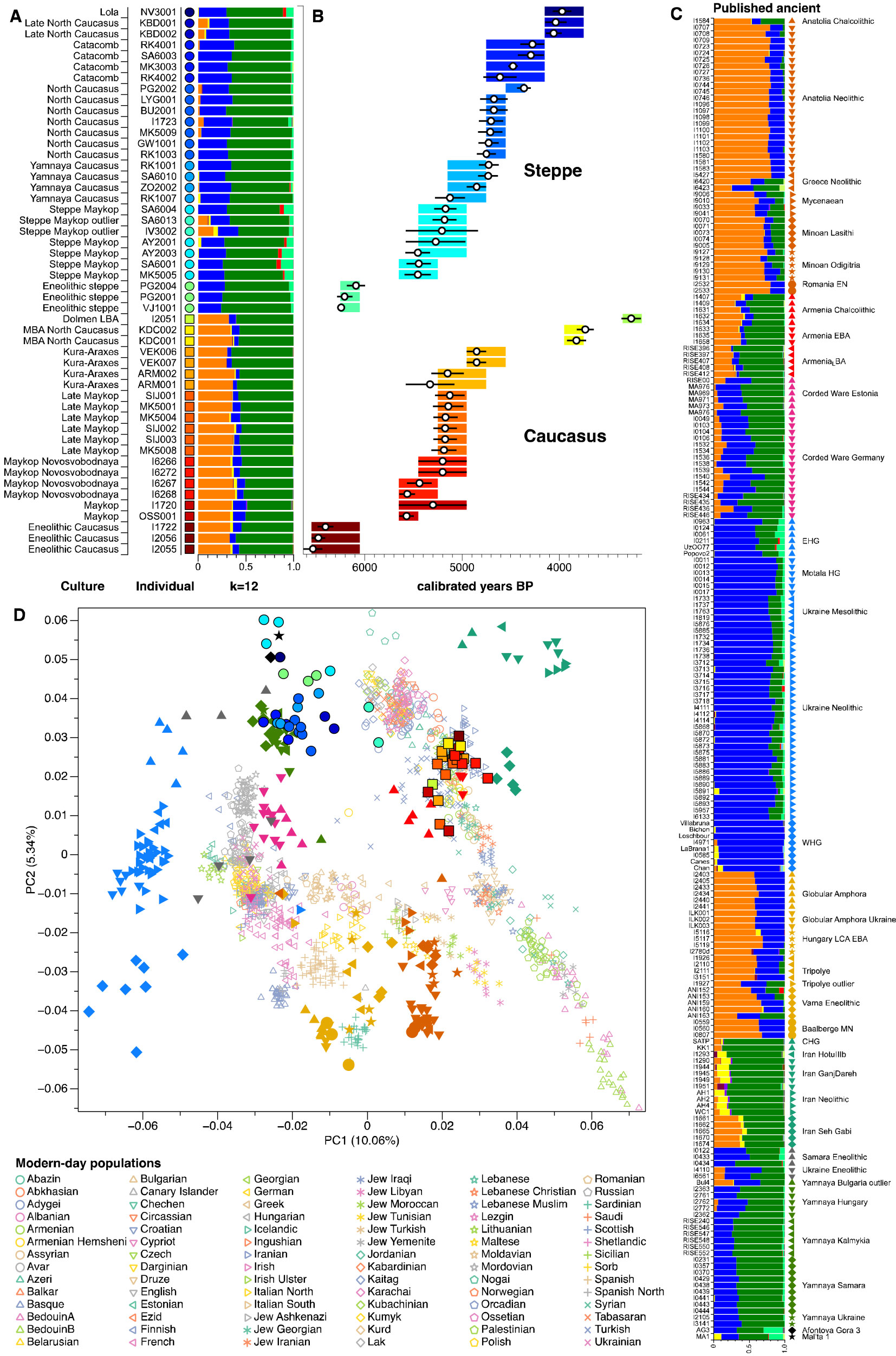
The origin of Q-M242 in Zvejnieki, like those of Lola (Q1a2-M25) and Steppe Maykop (Q1a2-M25) from Wang et al. (2018) are therefore most likely migrations throughout North Eurasia dated to the Palaeolithic.
As you might remember, the sample of haplogroup Q1a from Khvalynsk was the closest one (in the PCA, see above) to those we now know most likely represent one or more groups of the steppe north of the Caucasus, which were absorbed during the formation and expansion of Khvalynsk.
NOTE. In fact, the position of this early Khvalynsk sample in the PCA is near the Steppe Eneolithic cluster, in turn near ANE (with the Lola sample Q1a2-M25, circle in dark blue/violet above), and Steppe Maykop (which includes the other Q1a2-M25 sample).
It is often assumed that these populations absorbed in the Pontic-Caspian steppe were dominated by haplogroup J, due to the oldest representatives of CHG ancestry (Kotias Klde and Satsurblia).
However, it would not be surprising now to find out that (one or more of) these “CHG/ANE-rich” groups from the steppe (possibly the Kairshak culture in the North Caspian region) were in fact dominated by Q1-M25 subclades.
If this is the case, I don’t know where the proponents of the (south of the) Caucasus homeland will retreat to.
Related
- About Scepters, Horses, and War: on Khvalynsk migrants in the Caucasus and the Danube
- Steppe and Caucasus Eneolithic: the new keystones of the EHG-CHG-ANE ancestry in steppe groups
- The Caucasus a genetic and cultural barrier; Yamna dominated by R1b-M269; Yamna settlers in Hungary cluster with Yamna
- Consequences of Damgaard et al. 2018 (I): EHG ancestry in Maykop samples, and the potential Anatolian expansion routes
- Consequences of Damgaard et al. 2018 (II): The late Khvalynsk migration waves with R1b-L23 lineages
- No large-scale steppe migration into Anatolia; early Yamna migrations and MLBA brought LPIE dialects in Asia
- Early Indo-Iranian formed mainly by R1b-Z2103 and R1a-Z93, Corded Ware out of Late PIE-speaking migrations
- On the potential origin of Caucasus hunter-gatherer ancestry in Eneolithic steppe cultures
- Ancient human parallel lineages within North America contributed to a coastal expansion