Open access Ancient DNA shows domestic horses were introduced in the southern Caucasus and Anatolia during the Bronze Age by Guimaraes et al. Sci Adv. (2020) Vol. 6, no. 38, eabb0030.
Here is a good summary:
Our study of ancient equid remains from Anatolia and the southern Caucasus covering ~9000 years of the Holocene analyzed the dynamics over time of mitochondrial lineages and tested the hypothesis that Anatolia was a center of horse domestication. We were able to identify mitotypes characteristic of local Anatolian wild horses, which were regularly exploited in the early and middle Holocene. However, we identified a pattern of genetic change that does not reflect a gradual process involving the local population but rather a sudden appearance ~2000 BCE of nonlocal lineages that are still present in domestic horses. We also show that these imported horses exhibited coat colors that are absent in local wild horses before domestication. Moreover, continuation of Anatolian maternal lineage P into the Bronze Age implies some limited incorporation of local mares into domestic herds. These patterns of change indicate that domestic horses were introduced into Anatolia perhaps via the Caucasus region during the Bronze Age and provide a date for the beginning of the exploitation of domestic horses in Anatolia and Transcaucasia. They also argue against local independent domestication of the horse in this region. Our results strongly suggest that Anatolia was not a primary source for domestic horse lineages, but, as observed in other regions, local matrilines were incorporated into herds of imported domestic horses (…)

(Left) Evolution of mitochondrial haplotype diversity of horses in Anatolia and the southern Caucasus. (Right) Evolution of coat color genetic diversity in these two geographic regions in the same time ranges. The area of the circles is proportional to the number of individuals present in each category.
From the data itself, it is not difficult to see that the key period for the arrival of Proto-Anatolians (after ca. 4500 BC, and before ca. 3000 BC) is left without relevant information: the few Chalcolithic Çadir Höyük remains from Central Anatolia – first reported by Arbuckle – and the Lidar Höyük samples from Eastern Anatolia – without a clear radiocarbon date – leave us with no intermediate samples to check the ca. 2000 BC terminus ante quem that the authors describe.
If anything, it seems from the reported evolution of mtDNA and coat color – together with archaeology – that the Kura-Araxes expansion would fit best the emergence of those new horse breeds that swept through the Near East from the Southern Caucasus. Beyond that, it’s difficult to say anything else.
NOTE. For more on the potential relevance of the horse for Indo-Anatolians and the Kura-Araxes intrusion, see my recent posts on Anatolian origins and on Anatolian ancient DNA, and the section on Khvalynsk and Suvorovo-Novodanilovka.
This is a map with all published mtDNA to date:
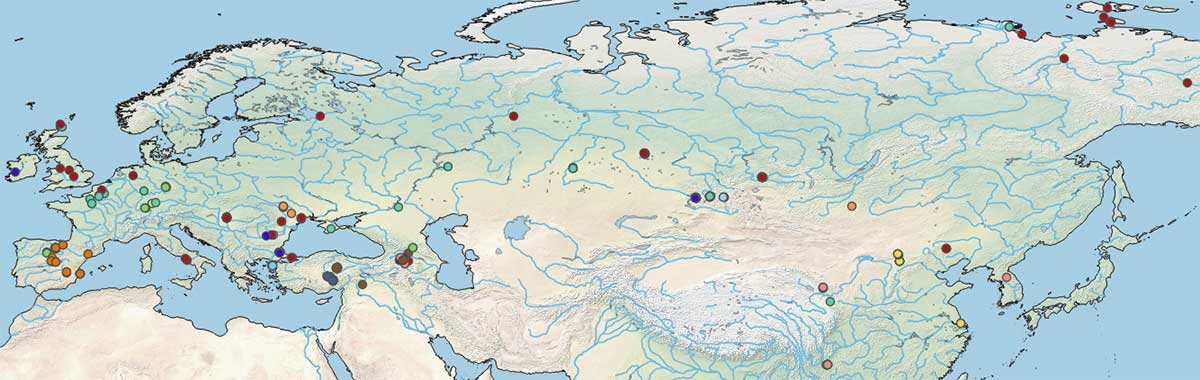
To make sense of its evolution, you can watch the video below:
Evolution of mtDNA adding the latest data on Anatolia and the Caucasus by Guimaraes et al. Sci. Adv. (2020) … 2/3 pic.twitter.com/367jVRiKbg
— Carlos Quiles (@cquilesc) September 19, 2020
After the hype of the abstract and the text, the reported data is obviously underwhelming. The information that is still lacking – a good chronological and geographical transect, nuclear DNA, and Y-HT – hinders a dismissal of a local or even primary independent domestication center in any of the regions sampled (southern Caucasus or Anatolia); or that some Chalcolithic or Early Bronze Age horses came from the Ancient DOM2 lineage; or that domestication emerged much earlier than 2000 BC and gradually instead of “suddenly”.
I’d agree that the authors are betting for the safest alternative in their interpretations, though, and that the reported data indirectly support what they say.
Nuclear DNA and Y chromosomal haplotypes
I was disappointed in the actual data, so I decided to take a look at what was already published, to see if I could make sense of anything else. This is a video with the evolution of nuclear DNA, Y chromosomal haplotypes, and sex – including also the nature of horses classified as ‘domestic’ or ‘wild’ based on their archaeological traits:
NOTE. For a previous discussion on the relevance of Fages et al. (2019), Gaunitz et al. (2018), Wutke et al. (2018), etc. see my latest post on how Yamnaya spread horse domesticates.
Evolution of horse domestication in nuDNA (Ancient DOM2, Iberian, Botai-Borly) and Y chrosomosome haplotypes, including other incomplete data on male vs. females and domestic vs. wild specimens (approximate century on bottom right) …1/3 pic.twitter.com/I5W3nLlack
— Carlos Quiles (@cquilesc) September 19, 2020
Below is a visual of the data divided into the same periods as the cultural maps of the Prehistory Atlas:

Future testing of available samples from the Middle Volga, Don and Dnieper might shed light on the genetic relationships of Early Eneolithic horses.

The Botai are the first sampled specimens from a lineage shared with the Borly individuals (and the modern Przewalski ones), but different from the Ancient DOM2.
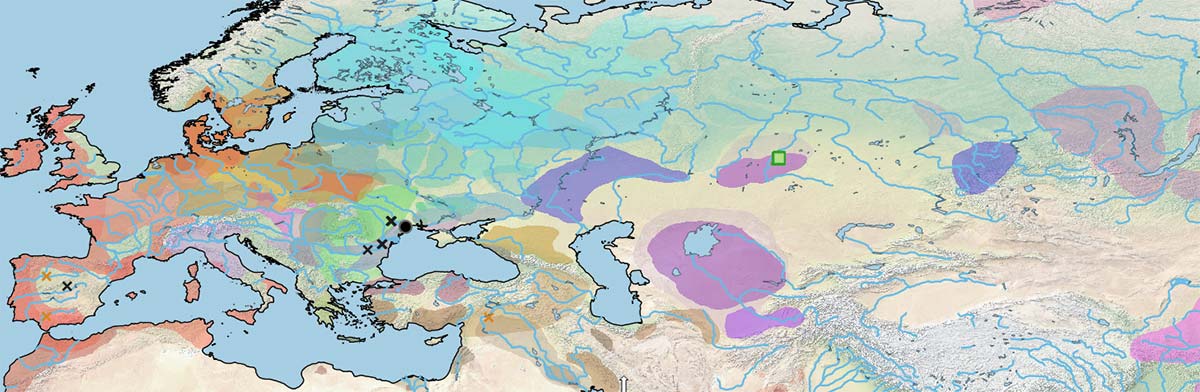
During the Final Eneolithic period, remains of wild horses keep popping up in the north Pontic area, at the same time as domestic ones (probably all Ancient DOM2-related) pop up in the Ural-Tobol interfluve, the Balkans, the Caucasus, and the Carpathian Basin, suggesting that hunter-gatherer populations from the north Pontic region still regarded horses as a source of meat. On the other hand, that is not incompatible with riding horses, as the Botai show.
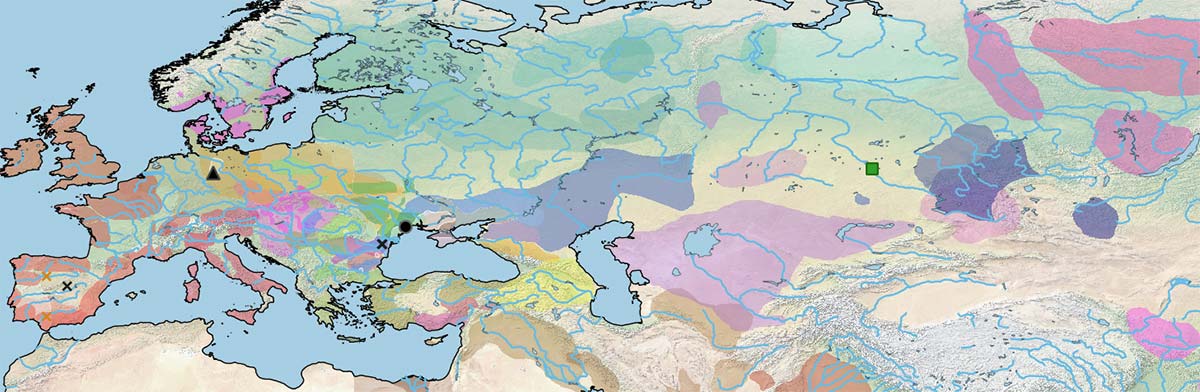
This is what Ludovic Orlando (2020) has to say about the horses sampled in South-East Europe:
Leopard-spotted horses seem to have lost their appeal after the early Bronze Age, a time when they were particularly abundant at the Turkish-Thrace site of Kirklareli-Kanligecit (2700– 2200 BCE), before they regained attention during the Iron Age. Interestingly, leopard spotting is associated with congenital night blindness, an incompletely dominant syndrome limiting vision in the dark in homozygous individuals. Leopard spotting is thus likely to be negatively selected in the wild, and the positive selection coefficients estimated at Kirklareli-Kanligecit suggest that the horse population analyzed was domesticated and not hunted. Whether these horses belonged to the DOM2 or the Botai-Borly4 lineage is presently unknown. However, the presence of the tobiano allele and the one causative for leopard spotting observed at Botai suggests that many different coat phenotypes contended at that site, which nicely complements archaeological arguments supporting the domestic status of these horses. Finally, even though ancient DNA data clearly established the rise of coat phenotypes with domestication, it is noteworthy that the dun, black, and leopard-spotted alleles have been documented in wild horses, including during the Upper Paleolithic.
The first known split in the phylogenetic tree of the modern domesticated horse lineage appears in a Hungarian specimen, Dunaujvaros_Duk2, from the late 3rd millennium BC (with a contribution ca. 38.6% from DOM2), and also in an Iron Age specimen from Els Vilars, north-east Iberia, of Pre-Iberian (likely Celtic) context. Both specimens had split from a branch older than the one found in Sintashta and succeeding Asian breeds, which supports the nature of West Yamnaya as horse riders, as described by Heyd (PNAS unpublished).
NOTE. As I said in the previous post on Fages et al. (2019), the “lineages” referred to when describing horse nuDNA concern the illustrative TreeMix phylogenetic trees that assume one migration pulse, which is obviously a simplification of the complex process of horse breeding in this and any other period…
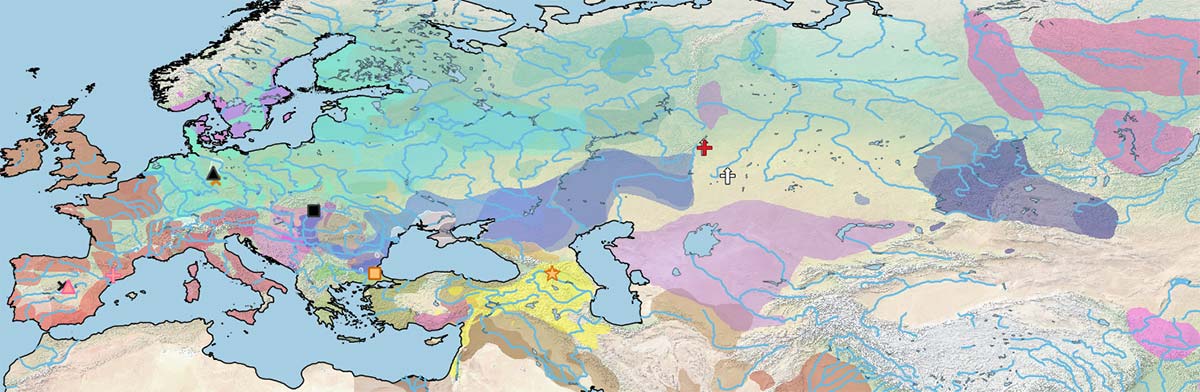
A female horse from Krasnokamenka, dated ca. 2983-1983 BC, is the oldest Ancient DOM2 specimen to appear in the maps. There is no information about its actual position in the phylogenetic tree, but based on its shared mtDNA with Sintashta-related specimens (Sintashta_NB44 and Halvai_KSH4) – continued in Bronze Age Asia and then spread into Iron Age Europe – I think it is safe to assume that it is from the same Sintashta nuDNA.
The authors only mention it by its mtDNA, and it is most likely not part of the previous Eneolithic Comb Ware-related period of the archaeological site. While it is possible that the true date lies at the end of the confidence interval (as with any wide radiocarbon date), it is interesting to remember how older dates from Sintashta materials are interpreted (from Anthony 2007):
The radiocarbon dates for both the cemeteries and the settlement at Sintashta were worryingly diverse, from about 2800-2700 BCE {4200+ 100 BP), for wood from grave 11 in the SM cemetery, to about 1800-1600 BCE (3340+60BP), for wood from grave 5 in the SII cemetery. Probably there was an older Poltavka component at Sintashta, as later was found at many other sites of the Sintashta type, accounting for the older dates.
Poltavka-culture herders had earlier occupied the northern steppe zone just where Sintashta appeared. The Poltavka culture was essentially a Volga-Ural continuation of the early Yamnaya horizon. Poltavka herding groups moved east into the Ural-Tobol steppes probably between 2800 and 2600 BCE. Poltavka decorative motifs on ceramics (vertical columns of chevrons) were very common on Sintashta pottery. A Poltavka kurgan cemetery (undated) stood on a low ridge 400 m south of the future site of Arkaim before that fortified settlement was built near the marshy bottom of the valley. The cemetery, Aleksandrovska IV, contained twnety-one small (10-20 m in diameter) kurgans, a relatively large Poltavka cemetery (figure 15.8). Six were excavated. All conformed to the typical Poltavka rite: a kurgan surrounded by a circular ditch, with a single grave with ledges, the body tightly contracted on the left or right side, lying on an organic mat, red ochre or white chalk by the head and occasionally around the whole body, with a pot or a flint tool or nothing. A few animal bones occasionally were dropped in the perimeter ditch. A Poltavka settlement was stratified beneath the Sintashta settlement of Kuisak, which is intriguing because Poltavka settlements, like Yamnaya settlements, are generally unknown. (…)
This is in full agreement with genomic finds of Yamnaya-related ancestry and hg. R1b-L23 among sampled Sintashta individuals up to the 2nd millennium BC.
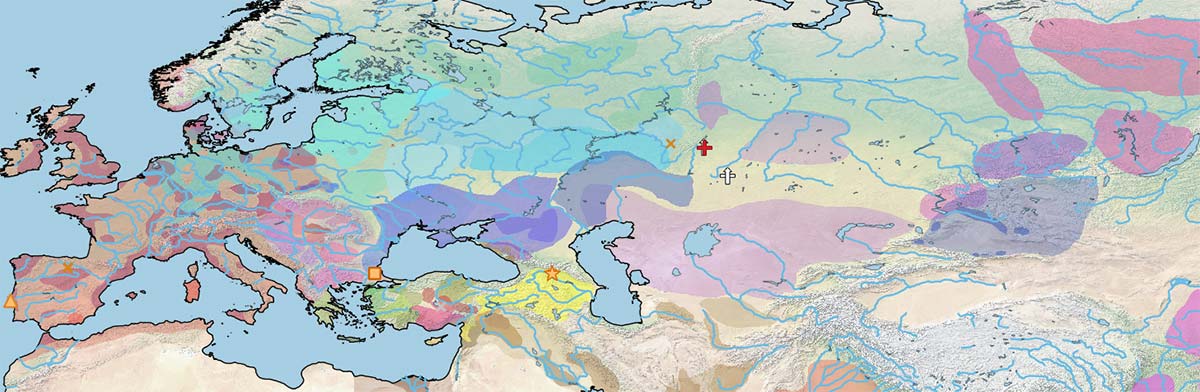
Only after the Bell Beaker period and the expansion of Sintashta-Potapovka and Abashevo-related peoples is the relevance of horseback riding patent in Europe and Asia.
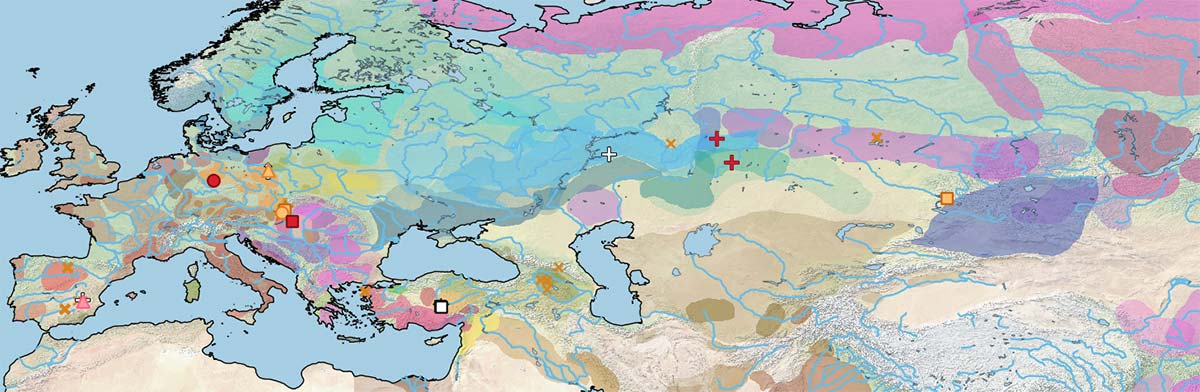
As for the increasing male representation since the Bronze Age reported in Fages et al. (2020), the maps show a good visual of this phenomenon.
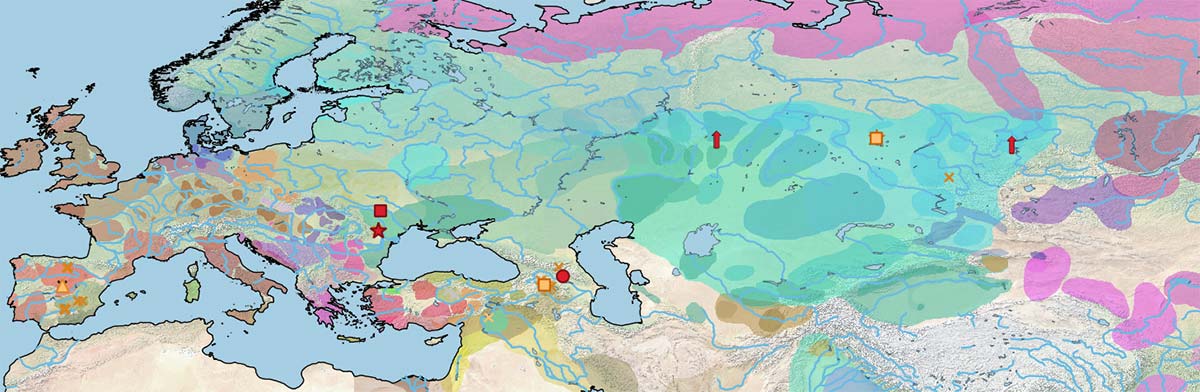
Looking carefully at the data century by century reveals that Y-HT-3 seems to have had some prevalence among the earliest DOM2 horses (both to the west and east of the steppes), while the other three haplotypes might have had a similar (slightly less relevant) distribution.
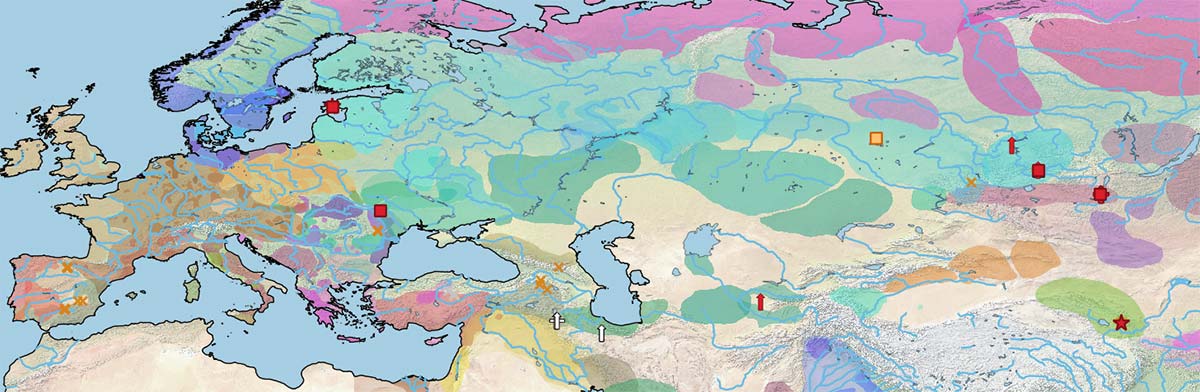
Y-HT-1 gradually dominated over the other three male lines, probably as a consequence of the different westward expansions of specialized horse riders from the Eurasian steppes, and later from Persia and during the Arab expansion.
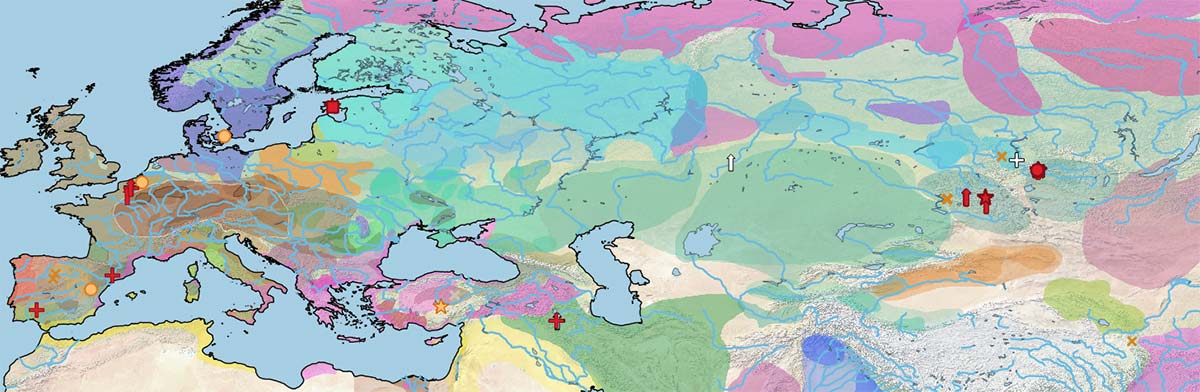
The often described replacement of stallions by Y chromosome haplotype Y-HT-1 is therefore not so marked as can be assumed from the maps in the original paper (Wutke et al. 2018), whereas mtDNA diversity becomes more limited that it is apparent in the maps above.
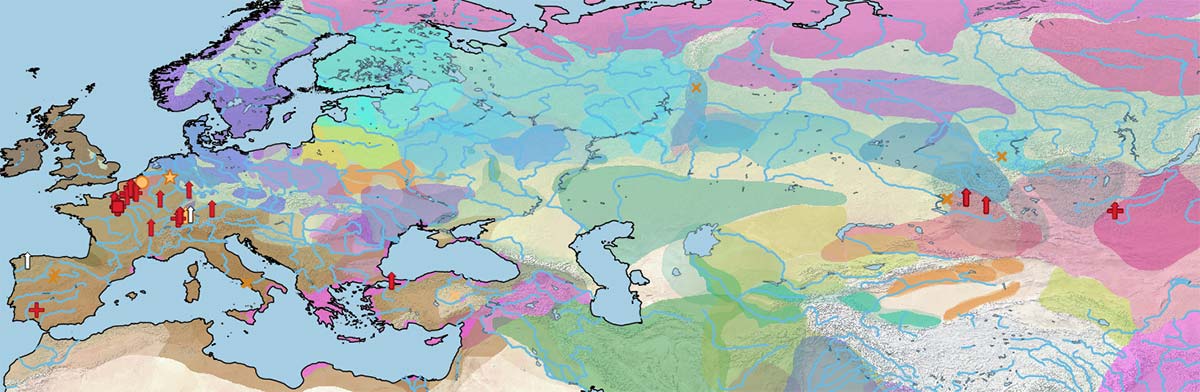
In any case, there is still a lack of proper evaluation of Y chromosome informative lines (such as in Felkel et al. 2019) to keep track of expanding male lines with enough precision.
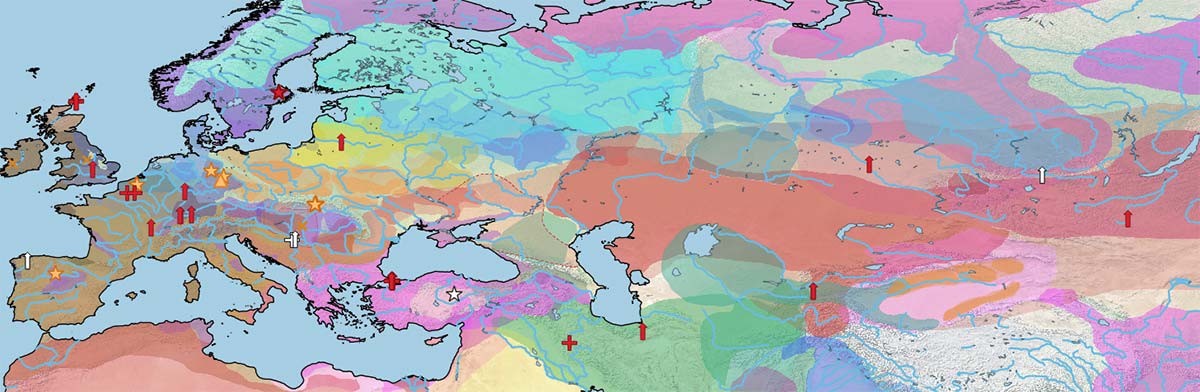
Conclusion
I have been reading some weird comments about this and other papers by academics, and I just want to lay out the basics as I understand them with regards to ancient horse DNA.
- When we talk about “domestic horses” in this kind of papers, the issue at hand is to ascertain the rough origin of the immediate ancestors of modern breeds, that were “initially” (that is, certainly by the Final Eneolithic period in the steppes) used mainly for horseback riding, but also later for traction and warfare.
- Not all domestic horses belong to this Ancient DOM2 lineage. Domestic horses were initially used as livestock, and early domesticates in Botai or (likely) Iberia show highly divergent ancestries. Maybe the domestication technique spread through cultural diffusion, but local horse breeds seem to have been prevalent during early domestication events, so multiple independent centers of domestication cannot be discarded.
- Not even ancient mounted horses were part of the same DOM2 lineages. The Botai lineage leading up to the Borly and (wild!) Przewalski horses are obvious examples of a highly divergent ancestry among horses that were ridden, and they were very close to the Pontic-Caspian steppes, where DOM2 probably emerged. Horse riding does look like something that could have been learnt through cultural diffusion in close contact.
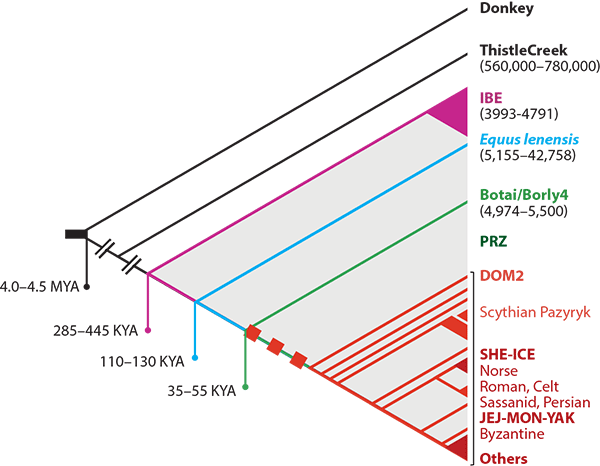
- Therefore, there probably were (and might appear in the ancient record) other domestic horses in the Pontic-Caspian steppes (and elsewhere) not belonging to the Ancient DOM2 lineage, that might have been ridden, too. That is an archaeological question, not a genetic one. There was probably very little (if anything at all) inherent to the earliest DOM2 as a better horse for domestication or riding, although many traits were consciously selected over time by breeders.
- The precise geographic origin of the DOM2 lineage does not determine anything precise about the Proto-Indo-European homeland. What is becoming clearer with each new wave of ancient DNA is that it was the horse-riding Yamnaya who spread them, likely stemming from an ancient tradition, but they might have selected to breed any kinds they deemed ‘better’ from any nearby region during the centuries they spread and remained in close contact.
- Hence, other ancient domestic horses from the Pontic-Caspian steppes (from, say, Samara, Dereivka, Khvalynsk, or Repin) might belong to some parallel branches (some “pre-Ancient DOM2” lineages), or to early diverging ones that survived in or around the steppes until the Yamnaya unified them all under the preferred selection.
In other words: the Ancient DOM2 ancestors could have been some specimens that might eventually be sampled among the Botai, Maykop, Dereivka, or – what is becoming more and more likely – Khvalynsk, but it would still be late Repin and later their offshoot Yamnaya the ultimate cause of the expansion of all modern domesticated horse breeds in West and East Eurasia.
See also
- Yamna the likely source of modern horse domesticates; the closest lineage, from East Bell Beakers
- Proto-Anatolians: from the Southern Caucasus or the Balkans?
- The genetic and cultural barrier of the Pontic-Caspian steppe – forest-steppe ecotone
- Mitogenomes suggest rapid expansion of domesticated horse before 3500 BC
- Origin of horse domestication likely on the North Caspian steppes
- Ancient DNA upends the horse family tree
- Domesticated horse population structure, selection, and mtDNA geographic patterns
- About Scepters, Horses, and War: on Khvalynsk migrants in the Caucasus and the Danube
- Steppe and Caucasus Eneolithic: the new keystones of the EHG-CHG-ANE ancestry in steppe groups
- Domestication spread probably via the North Pontic steppe to Khvalynsk… but not horse riding
- Differing modes of animal exploitation in North Pontic Eneolithic and Bronze Age Societies