New open access paper (in Chinese) A study of genetic diversity of three isolated populations in Xinjiang using Y-SNP, by liu et al. Acta Anthropologica Sinitica (2018)
Abstract:
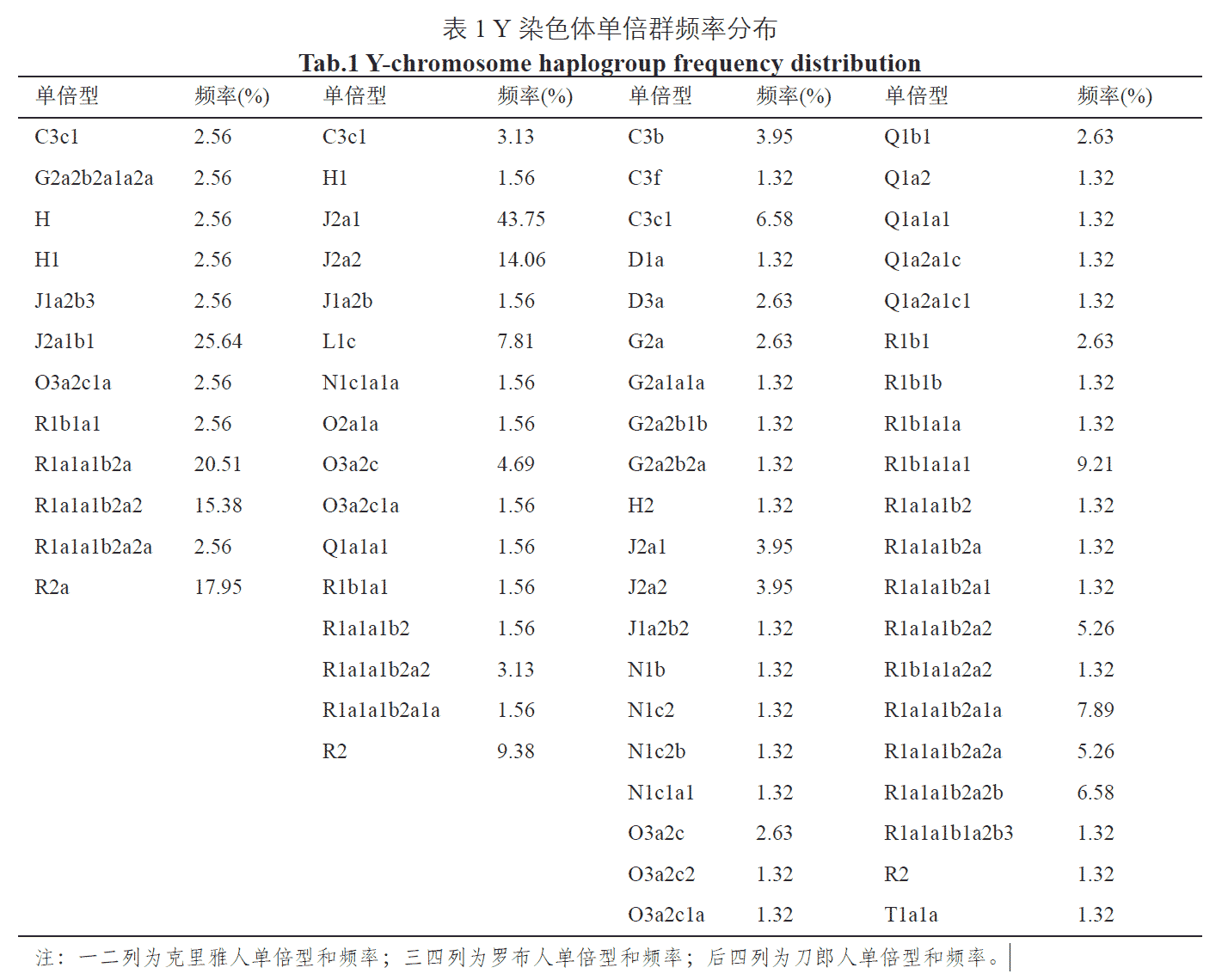
The Keriyan, Lopnur and Dolan peoples are isolated populations with sparse numbers living in the western border desert of our country. By sequencing and typing the complete Y-chromosome of 179 individuals in these three isolated populations, all mutations and SNPs in the Y-chromosome and their corresponding haplotypes were obtained. Types and frequencies of each haplotype were analyzed to investigate genetic diversity and genetic structure in the three isolated populations. The results showed that 12 haplogroups were detected in the Keriyan with high frequencies of the J2a1b1 (25.64%), R1a1a1b2a (20.51%), R2a (17.95%) and R1a1a1b2a2 (15.38%) groups. Sixteen haplogroups were noted in the Lopnur with the following frequencies: J2a1 (43.75%), J2a2 (14.06%), R2 (9.38%) and L1c (7.81%). Forty haplogroups were found in the Dolan, noting the following frequencies: R1b1a1a1 (9.21%), R1a1a1b2a1a (7.89%), R1a1a1b2a2b (6.58%) and C3c1 (6.58%). These data show that these three isolated populations have a closer genetic relationship with the Uygur, Mongolian and Sala peoples. In particular, there are no significant differences in haplotype and frequency between the three isolated populations and Uygur (f=0.833, p=0.367). In addition, the genetic haplotypes and frequencies in the three isolated populations showed marked Eurasian mixing illustrating typical characteristics of Central Asian populations.
My knowledge of written Chinese is almost zero, so here are some excerpts with the help of Google Translate:
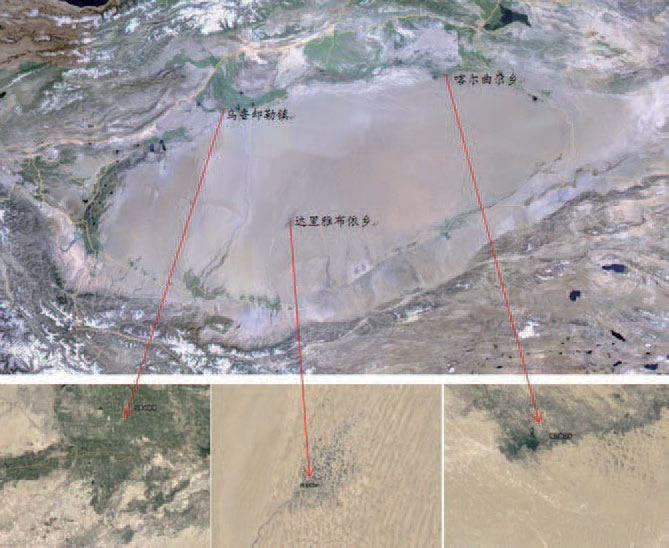
The source of 179 blood samples used in the study is shown in Figure 1. The Keriyan blood samples were collected from Dali Yabuyi Township, Yutian County (39 samples). The blood samples of the Lopnur people were collected from Kaerqu Township, Yuli County (64 cases); the blood samples of the Dolan people were collected from the town of Uluru, Awati County (76).
The composition and frequency of the Keriyan people’s haplogroup are closest to those of the Uighurs, and both Principal Component Analysis and Phylogenetic Tree Analysis show that their kinship is recent. We initially infer that the Keriyan are local desert indigenous people. They have a connection with the source of the Uighurs. Chen et al. [42] studied the patriarchal and maternal genetic analysis of the Keriyan people and found that they are not descendants of the Tibetan ethnic group in the West. The Keriyan people are a mixed group of Eastern and Western Europeans, which may originate from the local Vil group. Duan Ranhui [43] and other studies have shown that the nucleotide variability and average nucleotide differences in the Keriyan population are between the reported Eastern and Western populations. The phylogenetic tree also shows that the populations in Central Asia are between the continental lineage of the eastern population and the European lineage of the western population, and the genetic distance between the Keriyan and the Uighurs is the closest, indicating that they have a close relationship.
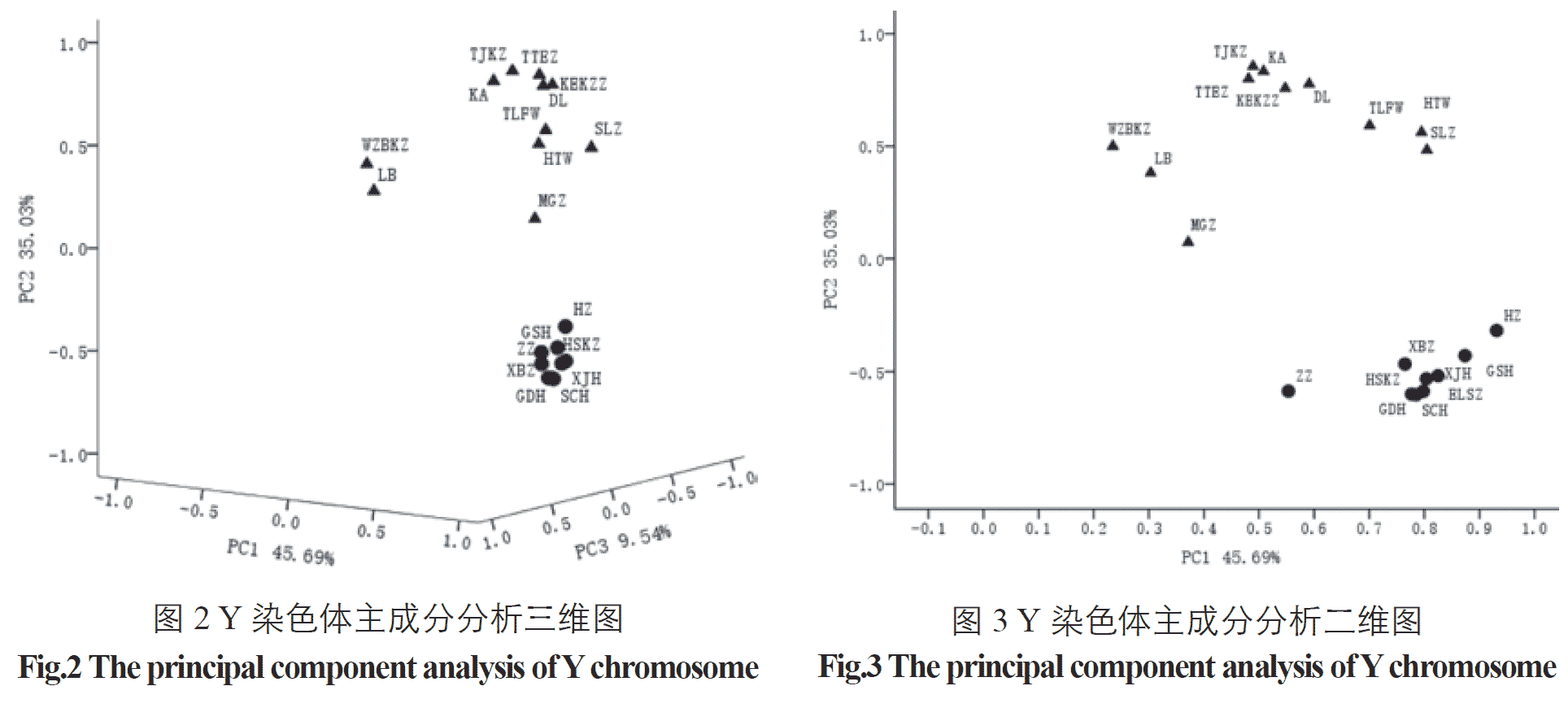
Regarding the origin of the Lopnur people, Purzhevski judged that it was a mixture of Mongolians and Aryans according to the physical characteristics of the Lopnur people. In 1934, the Sino-Swiss delegation discovered the famous burials of the ancient tombs in the Peacock River. After research, they were the indigenous people before the Loulan period; the researcher Yang Lan, a researcher at the Institute of Cultural Relics of the Chinese Academy of Social Sciences, said that the Lopnur people were descendants of the ancient “Landan survivors”. However, the Loulan people speaking an Indo-European language, and the Lopnur people speaking Uyghur languages contradict this; the historical materials of the Western Regions, “The Geography of the Western Regions” and “The Western Regions of the Ming Dynasty” record the Uighurs who lived in Cao Cao in the late 17th and early 18th centuries. Because of the occupation of the land by the Junggar nobles and their oppression, they fled. Some of them were forced to move to the Lop Nur area. There are many similar archaeological discoveries and historical records. We have no way to determine their accuracy, but they are at different times, and there is a great difference in what is heard in the same region. (…) The genetic characteristics of modern Lopnur people are the result of the long-term ethnic integration of Uyghurs, Mongols, and Europeans. This is also consistent with the similarity of the genetic structure of the Y chromosome of Lopnur in this study with the Uighurs and Mongolians. For example, the frequency of J haplogroup is as high as 59.37%, while J and its downstream sub-haplogroup are mainly distributed in western Europe, West Asia and Central Asia; the frequency of O, R haplogroup is close to that of Mongolians.
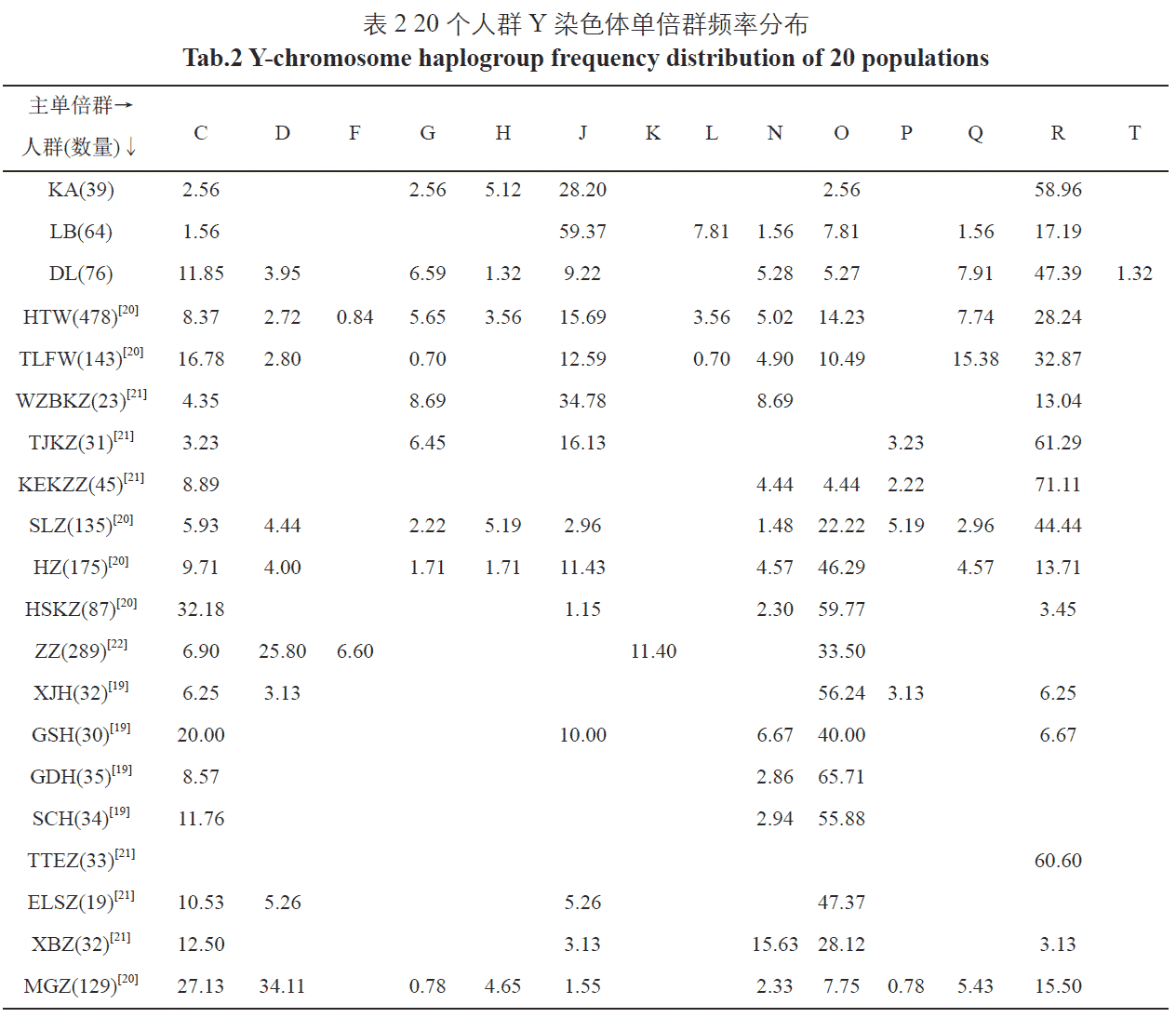
According to Ming History·Western Biography, the Mongolians originated from the Mobei Plateau and later ruled Asia and Eastern Europe. Mongolia was established, and large areas of southern Xinjiang and Central Asia were included. Later, due to the Mongolian king’s struggle for power, it fell into a long-term conflict. People of the land fled to avoid the war, and the uninhabited plain of the lower reaches of the Yarkant River naturally became a good place to live. People from all over the world gathered together and called themselves “Dura” and changed to “Dang Lang”. The long-term local Uyghur exchanges that entered the southern Mongolian monks and “Dura” were gradually assimilated [44]. According to the report, locals wore Mongolian clothes, especially women who still maintained a Mongolian face [45]. In 1976, the robes and waistbands found in the ancient time of the Daolang people in Awati County were very similar to those of the ancients. Dalang Muqam is an important part of Daolang culture. It is also a part of the Uyghur Twelve Muqam, and it retains the ancient Western culture, but it also contains a larger Mongolian culture and relics. The above historical records show that the Daolang people should appear in the Chagatai Khanate and be formed by the integration of Mongolian and Uighur ethnic groups. Through our research, we also found that the paternal haplotype of the Daolang people is contained in both Uygur and Mongolian, and the main haplogroups are the same, whereas the frequencies are different (see Table 3). The principal component analysis and the NJ analysis are also the same. It is very close to the Uyghur and the Mongolian people, which establishes new evidence for the “mixed theory” in molecular genetics.

If the nomenclature follows a recent ISOGG standard, it appears that:
The presence of exclusively R1a-Z93 subclades and the lack of R1b-M269 samples is compatible with the expansion of R1a-Z93 into the area with Proto-Tocharians, at the turn of the 3rd-2nd millennium BC, as suggested by the Xiaohe samples, supposedly R1a(xZ93).
Now that it is obvious from ancient DNA (as it was clear from linguistics) that Pre-Tocharians separated earlier than other Late PIE peoples, with the expansion of late Khvalynsk/Repin into the Altai, at the end of the 4th millennium, these prevalent R1a (probably Z93) samples may be showing a replacement of Pre-Tocharian Y-DNA with the Andronovo expansion already by 2000 BC.
Lacking proper assessment of ancient DNA from Proto-Tocharians, this potential early Y-DNA replacement is still speculative*. However, if that is the case, I wonder what the Copenhagen group will say when supporting this, but rejecting at the same time the more obvious Y-DNA replacement in East Yamna / Poltavka in the mid-3rd millennium with incoming Corded Ware-related peoples. I guess the invention of an Indo-Tocharian group may be near…
*NOTE. The presence of R1b-M269 among Proto-Tocharians, as well as the presence of R1b-M269 among Tarim Basin peoples in modern and ancient times is not yet fully discarded. The prevalence of R1a-Z93 may also be the sign of a more recent replacement by Iranian peoples, before the Mongolian and Turkic expansions that probably brought R1b(xM269).
Also, the presence of R1b (xM269) samples in east Asia strengthens the hypothesis of a back-migration of R1b-P297 subclades, from Northern Europe to the east, into the Lake Baikal area, during the Early Mesolithic, as found in the Botai samples and later also in Turkic populations – which are the most likely source of these subclades (and probably also of Q1a2 and N1c) in the region.
Related
- Yamna/Afanasevo elite males dominated by R1b-L23, Okunevo brings ancient Siberian/Asian population
- Steppe and Caucasus Eneolithic: the new keystones of the EHG-CHG-ANE ancestry in steppe groups
- North Asian mitogenomes hint at the arrival of pastoralists from West to East ca. 2800-1000 BC
- Consequences of Damgaard et al. 2018 (III): Proto-Finno-Ugric & Proto-Indo-Iranian in the North Caspian region
- Consequences of Damgaard et al. 2018 (II): The late Khvalynsk migration waves with R1b-L23 lineages
- Early Indo-Iranian formed mainly by R1b-Z2103 and R1a-Z93, Corded Ware out of Late PIE-speaking migrations
- The concept of “Outlier” in Human Ancestry (III): Late Neolithic samples from the Baltic region and origins of the Corded Ware culture
- Uralic as a Corded Ware substrate of Indo-Iranian, and loanwords in Finno-Ugric
- More evidence on the recent arrival of haplogroup N and gradual replacement of R1a lineages in North-Eastern Europe