New paper (behind paywall) by David Anthony, Archaeology, Genetics, and Language in the Steppes: A Comment on Bomhard, complementing in a favourable way Bomhard’s Caucasian substrate hypothesis in the current issue of the JIES.
NOTE. I have tried to access this issue for some days, but it’s just not indexed in my university library online service (ProQuest) yet. This particular paper is on Academia.edu, though, as are Bomhard’s papers on this issue in his site.
Interesting excerpts (emphasis mine):
Along the banks of the lower Volga many excavated hunting-fishing camp sites are dated 6200-4500 BC. They could be the source of CHG ancestry in the steppes. At about 6200 BC, when these camps were first established at Kair Shak III and Varfolomievka (42 and 28 on Figure 2), they hunted primarily saiga antelope around Dzhangar, south of the lower Volga, and almost exclusively onagers in the drier desert-steppes at Kair-Shak, north of the lower Volga. Farther north at the lower/middle Volga ecotone, at sites such as Varfolomievka and Oroshaemoe hunter-fishers who made pottery similar to that at Kair-Shak hunted onagers and saiga antelope in the desert-steppe, horses in the steppe, and aurochs in the riverine forests. Finally, in the Volga steppes north of Saratov and near Samara, hunter-fishers who made a different kind of pottery (Samara type) and hunted wild horses and red deer definitely were EHG. A Samara hunter-gatherer of this era buried at Lebyazhinka IV, dated 5600-5500 BC, was one of the first named examples of the EHG genetic type (Haak et al. 2015). This individual, like others from the same region, had no or very little CHG ancestry. The CHG mating network had not yet reached Samara by 5500 BC.
But before 4500 BC, CHG ancestry appeared among the EHG hunter-fishers in the middle Volga steppes from Samara to Saratov, at the same time that domesticated cattle and sheep-goats appeared. The Reich lab now has whole-genome aDNA data from more than 30 individuals from three Eneolithic cemeteries in the Volga steppes between the cities of Saratov and Samara (Khlopkov Bugor, Khvalynsk, and Ekaterinovka), all dated around the middle of the fifth millennium BC. Many dates from human bone are older, even before 5000 BC, but they are affected by strong reservoir effects, derived from a diet rich in fish, making them appear too old (Shishlina et al 2009), so the dates I use here accord with published and unpublished dates from a few dated animal bones (not fish-eaters) in graves.
Only three individuals from Khvalynsk are published, and they were first published in a report that did not mention the site in the text (Mathieson et al. 2015), so they went largely unnoticed. Nevertheless, they are crucial for understanding the evolution of the Yamnaya mating network in the steppes. They were mentioned briefly in Damgaard et al (2018) but were not graphed. They were re-analyzed and their admixture components were illustrated in a bar graph in Wang et al (2018: figure 2c), but they are not the principal focus of any published study. All of the authors who examined them agreed that these three Khvalynsk individuals, dated about 4500 BC, showed EHG ancestry admixed substantially with CHG, and not a trace of Anatolian Farmer ancestry, so the CHG was a Hotu-Cave or Kotias-Cave type of un-admixed CHG. The proportion of CHG in the Wang et al. (2018) bar graphs is about 20-30% in two individuals, substantially less CHG than in Yamnaya; but the third Khvalynsk individual had more than 50% CHG, like Yamnaya. The ca. 30 additional unpublished individuals from three middle Volga Eneolithic cemeteries, including Khvalynsk, preliminarily show the same admixed EHG/CHG ancestry in varying proportions. Most of the males belonged to Y-chromosome haplogroup R1b1a, like almost all Yamnaya males, but Khvalynsk also had some minority Y-chromosome haplogroups (R1a, Q1a, J, I2a2) that do not appear or appear only rarely (I2a2) in Yamnaya graves.
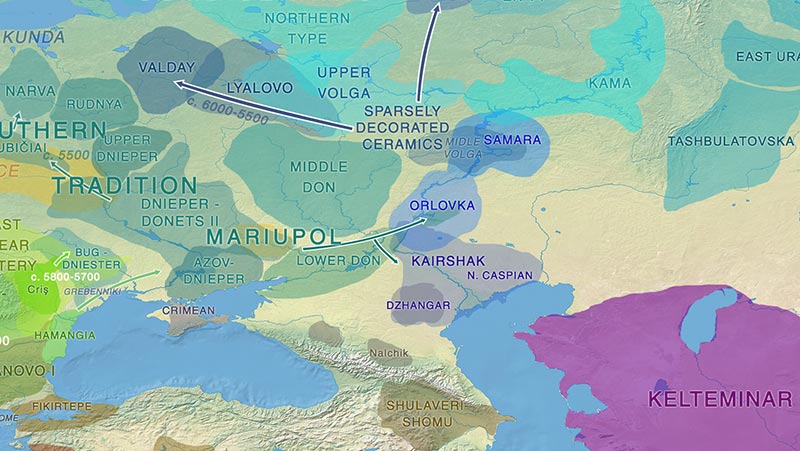
Pontic-Caspian steppe and neighbouring groups in the Neolithic. See full map.
Wang et al. (2018) discovered that this middle Volga mating network extended down to the North Caucasian steppes, where at cemeteries such as Progress-2 and Vonyuchka, dated 4300 BC, the same Khvalynsk-type ancestry appeared, an admixture of CHG and EHG with no Anatolian Farmer ancestry, with steppe-derived Y-chromosome haplogroup R1b. These three individuals in the North Caucasus steppes had higher proportions of CHG, overlapping Yamnaya. Without any doubt, a CHG population that was not admixed with Anatolian Farmers mated with EHG populations in the Volga steppes and in the North Caucasus steppes before 4500 BC. We can refer to this admixture as pre-Yamnaya, because it makes the best currently known genetic ancestor for EHG/CHG R1b Yamnaya genomes. The Progress-2 individuals from North Caucasus steppe graves lived not far from the pre-Maikop farmers of the Belaya valley, but they did not exchange mates, according to their DNA.
The hunter-fisher camps that first appeared on the lower Volga around 6200 BC could represent the migration northward of un-admixed CHG hunter-fishers from the steppe parts of the southeastern Caucasus, a speculation that awaits confirmation from aDNA. After 5000 BC domesticated animals appeared in these same sites in the lower Volga, and in new ones, and in grave sacrifices at Khvalynsk and Ekaterinovka. CHG genes and domesticated animals flowed north up the Volga, and EHG genes flowed south into the North Caucasus steppes, and the two components became admixed. After approximately 4500 BC the Khvalynsk archaeological culture united the lower and middle Volga archaeological sites into one variable archaeological culture that kept domesticated sheep, goats, and cattle (and possibly horses). In my estimation, Khvalynsk might represent the oldest phase of PIE.
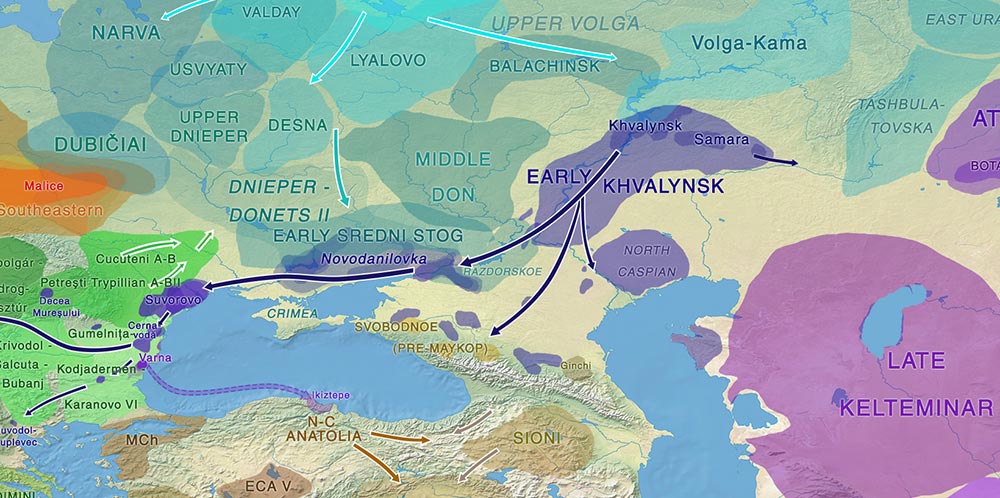
Pontic-Caspian steppe and neighbouring groups in the Early Eneolithic. See full map.
Anatolian Farmer ancestry and Yamnaya origins
The Eneolithic Volga-North Caucasus mating network (Khvalynsk/Progress-2 type) exhibited EHG/CHG admixtures and Y-chromosome haplogroups similar to Yamnaya, but without Yamnaya’s additional Anatolian Farmer ancestry. (…)
Like the Mesolithic and Neolithic populations here, the Eneolithic populations of Dnieper-Donets II type seem to have limited their mating network to the rich, strategic region they occupied, centered on the Rapids. The absence of CHG shows that they did not mate frequently if at all with the people of the Volga steppes, a surprising but undeniable discovery. Archaeologists have seen connections in ornament types and in some details of funeral ritual between Dnieper-Donets cemeteries of the Mariupol-Nikol’skoe type and cemeteries in the middle Volga steppes such as Khvalynsk and S’yez’zhe (Vasiliev 1981:122-123). Also their cranio-facial types were judged to be similar (Bogdanov and Khokhlov 2012:212). So it it surprising that their aDNA does not indicate any genetic admixture with Khvalynsk or Progress-2. Also, neither they nor the Volga steppe Eneolithic populations showed any Anatolian Farmer ancestry. (…)
All three of the steppe-admixed exceptions were from the Varna region (Mathieson et al. 2018). One of them was the famous “golden man’ at Varna (Krause et al. 2016), Grave 43, whose steppe ancestry was the most doubtful of the three. If he had steppe ancestry, it was sufficiently distant (five+ generations before him) that he was not a statistically significant outlier, but he was displaced in the steppe direction, away from the central values of the majority of typical Anatolian Farmers at Varna and elsewhere. The other two, at Varna (grave 158, a 5-7-year-old girl) and Smyadovo (grave 29, a male 20-25 years old), were statistically significant outliers who had recent steppe ancestry (consistent with grandparents or great-grandparents) of the EHG/CHG Khvalynsk/Progress-2 type, not of the Dnieper Rapids EHG/WHG type.
(…) I believe that the Suvorovo-Cernavoda I movement into the lower Danube valley and the Balkans about 4300 BC separated early PIE-speakers (pre-Anatolian) from the steppe population that stayed behind in the steppes and that later developed into late PIE and Yamnaya.
This archaeological transition marked the breakdown of the mating barrier between steppe and Anatolian Farmer mating networks. After this 4300-4200 BC event, Anatolian Farmer ancestry began to pop up in the steppes. The currently oldest sample with Anatolian Farmer ancestry in the steppes in an individual at Aleksandriya, a Sredni Stog cemetery on the Donets in eastern Ukraine. Sredni Stog has often been discussed as a possible Yamnaya ancestor in Ukraine (Anthony 2007: 239- 254). The single published grave is dated about 4000 BC (4045– 3974 calBC/ 5215±20 BP/ PSUAMS-2832) and shows 20% Anatolian Farmer ancestry and 80% Khvalynsk-type steppe ancestry (CHG&EHG). His Y-chromosome haplogroup was R1a-Z93, similar to the later Sintashta culture and to South Asian Indo-Aryans, and he is the earliest known sample to show the genetic adaptation to lactase persistence (I3910-T). Another pre-Yamnaya grave with Anatolian Farmer ancestry was analyzed from the Dnieper valley at Dereivka, dated 3600-3400 BC (grave 73, 3634–3377 calBC/ 4725±25 BP/ UCIAMS-186349). She also had 20% Anatolian Farmer ancestry, but she showed less CHG than Aleksandriya and more Dereivka-1 ancestry, not surprising for a Dnieper valley sample, but also showing that the old fifth-millennium-type EHG/WHG Dnieper ancestry survived into the fourth millennium BC in the Dnieper valley (Mathieson et al. 2018).
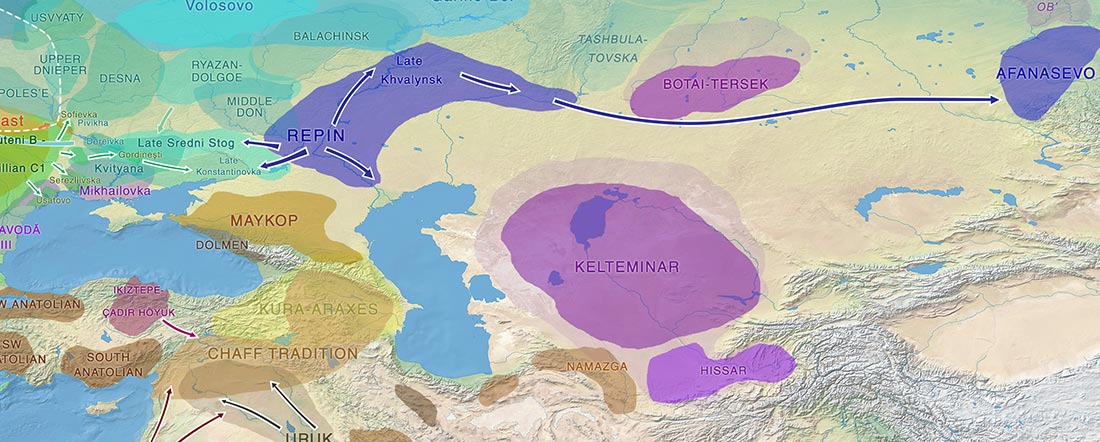
Pontic-Caspian steppe and neighbouring groups in the Late Eneolithic. See full map.
Probably, late PIE (Yamnaya) evolved in the same part of the steppes—the Volga-Caucasus steppes between the lower Don, the lower and middle Volga, and the North Caucasus piedmont—where early PIE evolved, and where appropriate EHG/CHG admixtures and Y-chromosome haplogroups were seen already in the Eneolithic (without Anatolian Farmer). There have always been archaeologists who argued for an origin of Yamnaya in the Volga steppes, including Gimbutas (1963), Merpert (1974), and recently Morgunova (2014), who argued that this was where Repin-type ceramics, an important early Yamnaya pottery type, first appeared in dated contexts before Yamnaya, about 3600 BC. The genetic evidence is consistent with Yamnaya EHG/CHG origins in the Volga-Caucasus steppes. Also, if contact with the Maikop culture was a fundamental cause of the innovations in transport and metallurgy that defined the Yamnaya culture, then the lower Don-North Caucasus-lower Volga steppes, closest to the North Caucasus, would be where the earliest phase is expected.
I would still guess that the Darkveti-Meshoko culture and its descendant Maikop culture established the linguistic ancestor of the Northwest Caucasian languages in approximately the region where they remained. I also accept the general consensus that the appearance of the hierarchical Maikop culture about 3600 BC had profound effects on pre-Yamnaya and early Yamnaya steppe cultures. Yamnaya metallurgy borrowed from the Maikop culture two-sided molds, tanged daggers, cast shaft hole axes with a single blade, and arsenical copper. Wheeled vehicles might have entered the steppes through Maikop, revolutionizing steppe economies and making Yamnaya pastoral nomadism possible after 3300 BC.
For those who still hoped that Proto-Indo-Europeans of Yamnaya/Afanasievo ancestry from the Don-Volga region were associated with the expansion of hg. R1a-M417, in a sort of mythical “R1-rich” Indo-European society, it seems this is going to be yet another prediction based on ancestry magic that goes wrong.
Proto-Indo-Europeans were, however, associated with other subclades beyond R1b-M269, probably (as I wrote recently) R1b-V1636, I2a-L699, Q1a-M25, and R1a-YP1272, but also interestingly some J subclade, so let’s see what surprises the new study on Khvalynsk and Yamnaya settlers from the Carpathian Basin brings…
On the bright side, it is indirectly confirmed that late Sredni Stog formed part of the neighbouring Corded Ware-like populations of ca. 20-30%+ Anatolian farmer ancestry that gave Yamnaya its share (ca. 6-10%), relative to the comparatively unmixed Khvalynsk and late Repin population (as shown by Afanasevo).
In this steppe mating network that opened up after the Khvalynsk expansion, the increasing admixture of Anatolian farmer-related ancestry in Yamnaya from east (ca. 2-10%) to west (ca. 6-15%) points to an exogamy of late Repin males in their western/south-western regions with populations around the Don River basin and beyond (and endogamy within the Yamnaya community), in an evolution relevant for language expansions and language contacts during the Late Eneolithic.
NOTE. “Mating network” is my new preferred term for “ancestry”. Also great to see scholars finally talk about “Pre-Yamnaya” ancestry, which – combined with the distinction of Yamnaya from Corded Ware ancestry – will no doubt help differentiate fine-scale population movements of steppe- and forest-steppe-related populations.
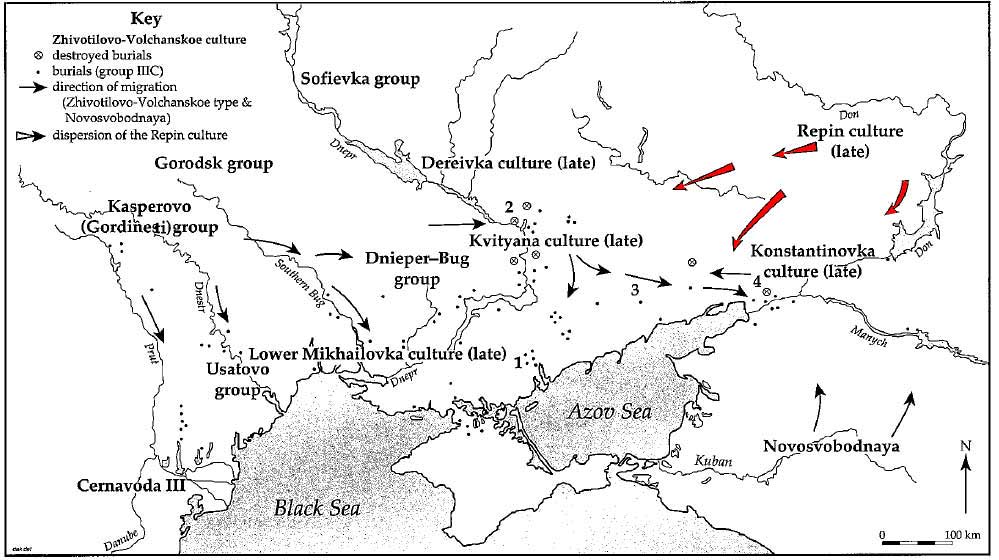
Modified from Rassamakin (1999), adding red color to Repin expansion. The system of the latest Eneolithic Pointic cultures and the sites of the Zhivotilovo-Volchanskoe type: 1) Volchanskoe; 2) Zhivotilovka; 3) Vishnevatoe; 4) Koisug.
On the other hand, it would also be interesting to read a discussion of how this Volga homeland of Middle PIE and Don-Volga-Ural homeland of Late PIE would be reconciled with the known continuous contacts of Uralic with Middle and Late PIE (see here) to locate the most likely Proto-Uralic homeland.
Especially because Corded Ware fully replaced all sub-Neolithic groups to the north and east of Khvalynsk/Yamnaya, like Volosovo, so no other population neighbouring Middle and Late Proto-Indo-Europeans survived into the Bronze Age…
It is good practice to be registered
and logged in to comment.
Please keep the discussion of this post on topic. Civilized discussion. Academic tone.
For other topics, use the forums instead.