The preprint by Jeong et al. (2018) has been published: The genetic history of admixture across inner Eurasia Nature Ecol. Evol. (2019).
Interesting excerpts, referring mainly to Uralic peoples (emphasis mine):
A model-based clustering analysis using ADMIXTURE shows a similar pattern (Fig. 2b and Supplementary Fig. 3). Overall, the proportions of ancestry components associated with Eastern or Western Eurasians are well correlated with longitude in inner Eurasians (Fig. 3). Notable outliers include known historical migrants such as Kalmyks, Nogais and Dungans. The Uralic- and Yeniseian-speaking populations, as well as Russians from multiple locations, derive most of their Eastern Eurasian ancestry from a component most enriched in Nganasans, while Turkic/Mongolic speakers have this component together with another component most enriched in populations from the Russian Far East, such as Ulchi and Nivkh (Supplementary Fig. 3). Turkic/Mongolic speakers comprising the bottom-most cline have a distinct Western Eurasian ancestry profile: they have a high proportion of a component most enriched in Mesolithic Caucasus hunter-gatherers and Neolithic Iranians and frequently harbour another component enriched in present-day South Asians (Supplementary Fig. 4). Based on the PCA and ADMIXTURE results, we heuristically assigned inner Eurasians to three clines: the ‘forest-tundra’ cline includes Russians and all Uralic and Yeniseian speakers; the ‘steppe-forest’ cline includes Turkic- and Mongolic-speaking populations from the Volga and Altai–Sayan regions and Southern Siberia; and the ‘southern steppe’ cline includes the rest of the populations.
For the forest-tundra populations, the Nganasan + Srubnaya model is adequate only for the two Volga region populations, Udmurts and Besermyans (Fig. 5 and Supplementary Table 8).
For the other populations west of the Urals, six from the northeastern corner of Europe are modelled with additional Mesolithic Western European hunter-gatherer (WHG) contribution (8.2–11.4%; Supplementary Table 8), while the rest need both WHG and early Neolithic European farmers (LBK_EN; Supplementary Table 2). Nganasan-related ancestry substantially contributes to their gene pools and cannot be removed from the model without a significant decrease in the model fit (4.1–29.0% contribution; χ2 P ≤ 1.68 × 10−5; Supplementary Table 8).
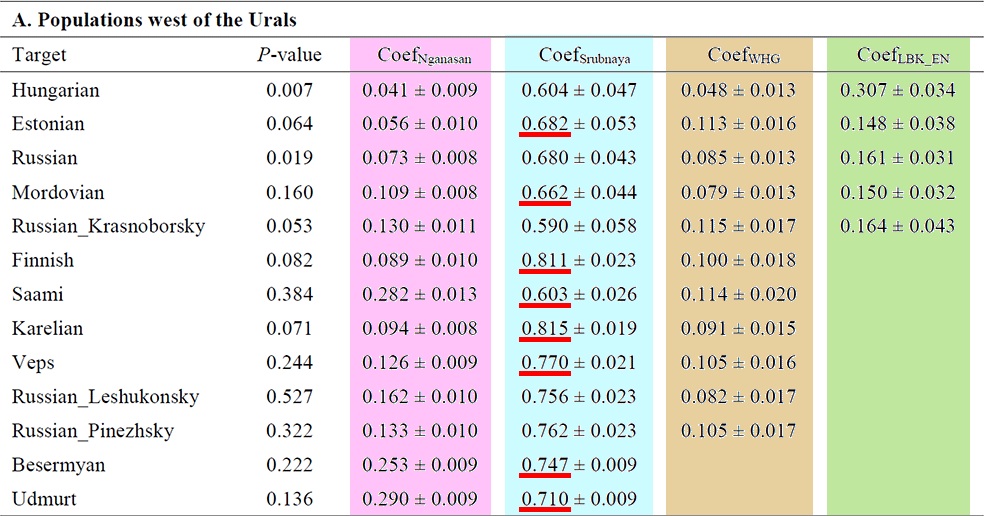
NOTE. It doesn’t seem like Hungarians can be easily modelled with Nganasan ancestry, though…
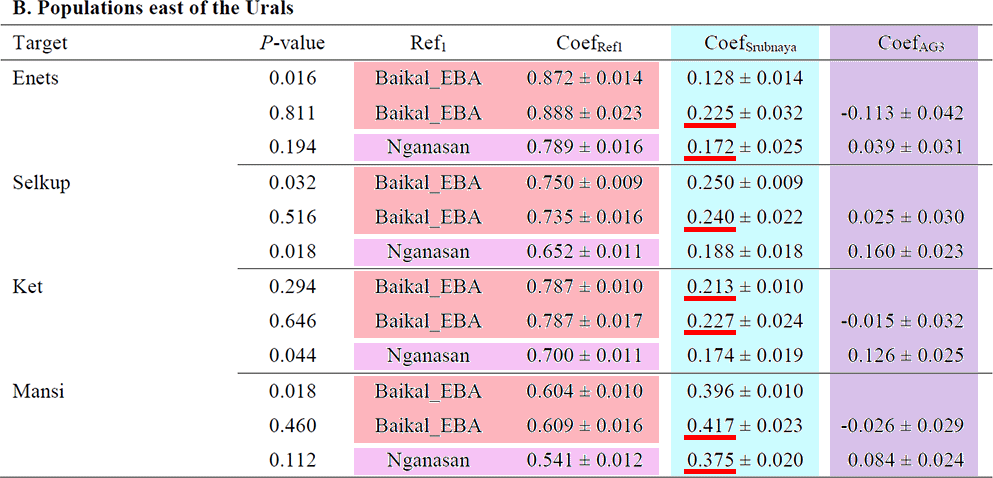
For the 4 populations east of the Urals (Enets, Selkups, Kets and Mansi), for which the above models are not adequate, Nganasan + Srubnaya + AG3 provides a good fit (χ2 P ≥ 0.018; Fig. 5 and Supplementary Table 8). Using early Bronze Age populations from the Baikal Lake region (‘Baikal_EBA’; Supplementary Table 2) as a reference instead of Nganasan, the two-way model of Baikal_EBA + Srubnaya provides a reasonable fit (χ2 P ≥ 0.016; Supplementary Table 8) and the three-way model of Baikal_EBA + Srubnaya + AG3 is adequate but with negative AG3 contribution for Enets and Mansi (χ2 P ≥ 0.460; Supplementary Table 8).
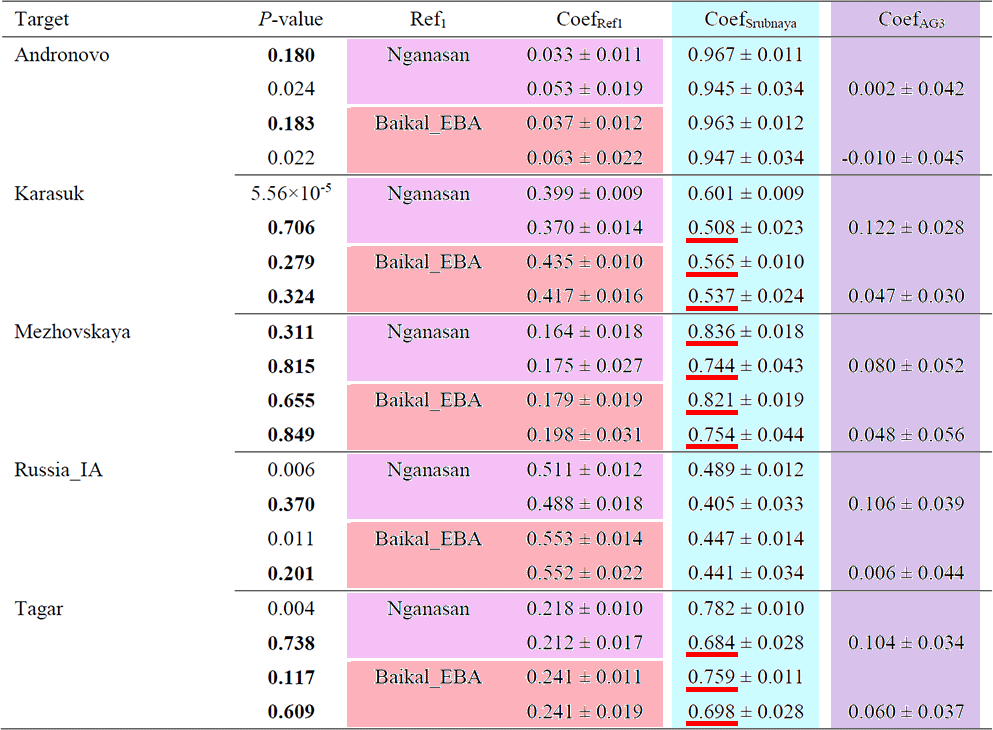
Bronze/Iron Age populations from Southern Siberia also show a similar ancestry composition with high ANE affinity (Supplementary Table 9). The additional ANE contribution beyond the Nganasan + Srubnaya model suggests a legacy from ANE-ancestry-rich clines before the Late Bronze Age.
Lara M. Cassidy comments the results of the study in A steppe in the right direction (you can read it here):
Even among the earliest available inner Eurasian genomes, east–west connectivity is evident. These, too, form a longitudinal cline, characterized by the easterly increase of a distinct ancestry, labelled Ancient North Eurasian (ANE), lowest in western European hunter-gatherers (WHG) and highest in Palaeolithic Siberians from the Baikal region. Flow-through from this ANE cline is seen in steppe populations until at least the Bronze Age, including the world’s earliest known horse herders — the Botai. However, this is eroded over time by migration from west and east, following agricultural adoption on the continental peripheries (Fig. 1b,c).
Strikingly, Jeong et al. model the modern upper steppe cline as a simple two-way mixture between western Late Bronze Age herders and Northeast Asians (Fig. 1c), with no detectable residue from the older ANE cline. They propose modern steppe peoples were established mainly through migrations post-dating the Bronze Age, a sequence for which has been recently outlined using ancient genomes. In contrast, they confirm a substantial ANE legacy in modern Siberians of the northernmost cline, a pattern mirrored in excesses of WHG ancestry west of the Urals (Fig. 1b). This marks the inhospitable biome as a reservoir for older lineages, an indication that longstanding barriers to latitudinal movement may indeed be at work, reducing the penetrance of gene flows further south along the steppe.
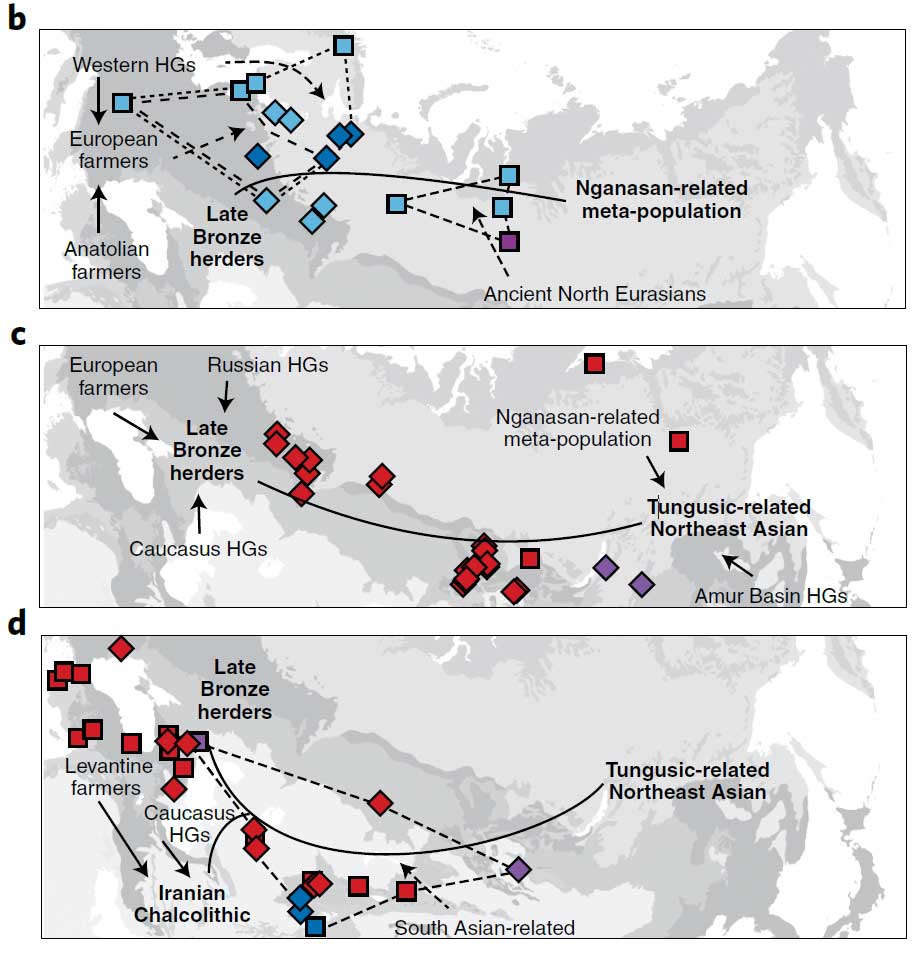
Given the findings as reported in the paper, I think it should be much easier to describe different subclines in the “northernmost cline” than in the much more recent “Turkic/Mongolic cline”, which is nevertheless subdivided in this paper in two clines. As an example, there are at least two obvious clines with “Nganasan-related meta-populations” among Uralians, which converge in a common Steppe MLBA (i.e. Corded Ware) ancestry – one with Palaeo-Laplandic peoples, and another one with different Palaeo-Siberian populations:
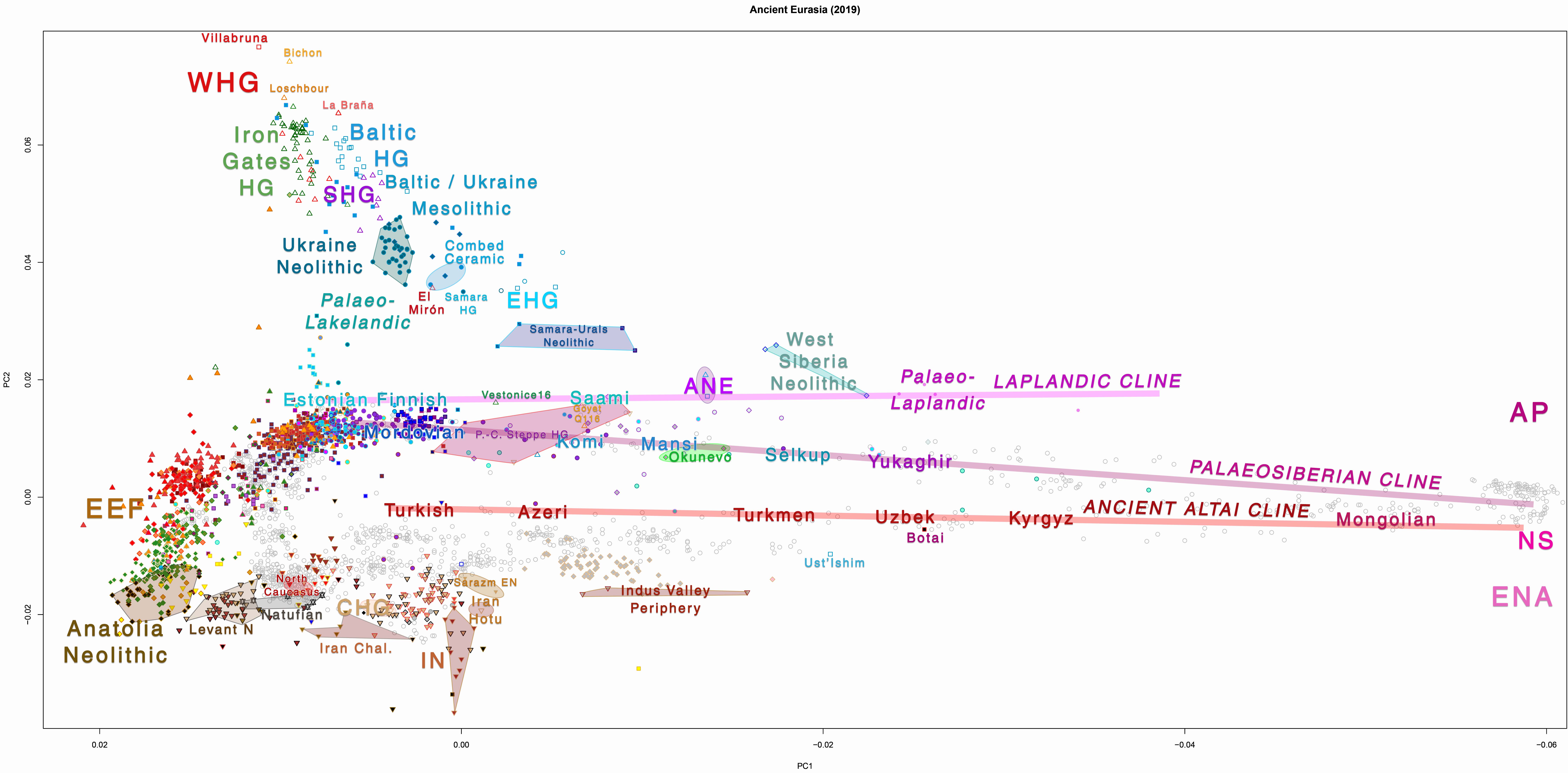
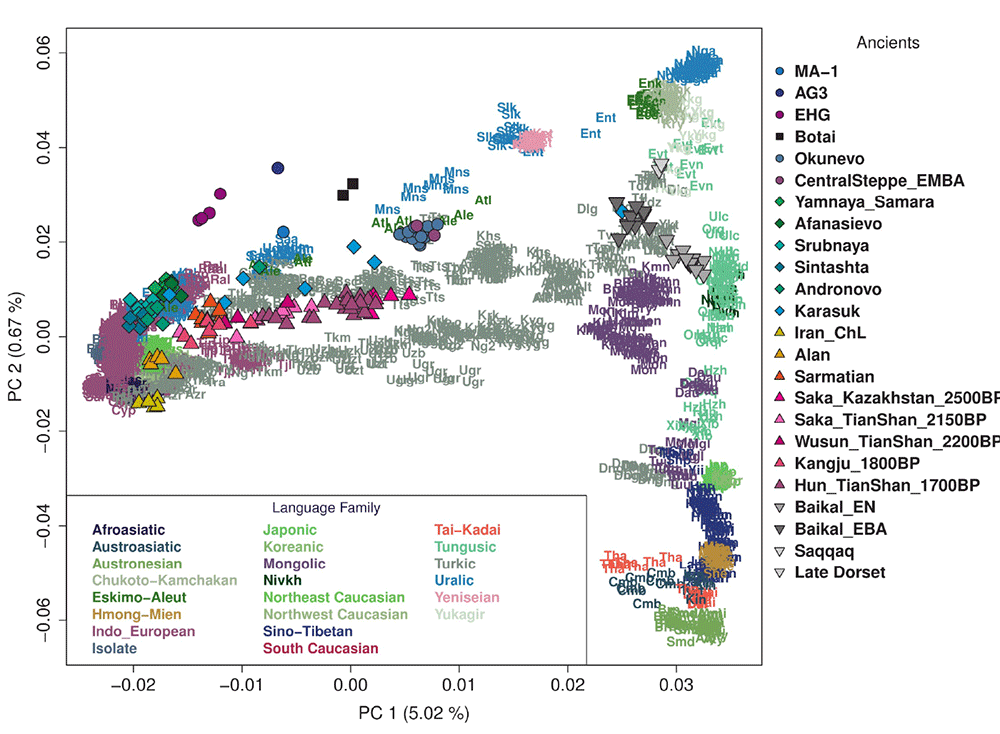
The inclusion of certain Eurasian groups (or lack thereof) in the PCA doesn’t help to distinguish these subclines visually, and I guess the tiny “Naganasan-related” ancestral components found in some western populations (e.g. the famous ~5% among Estonians) probably don’t lend themselves easily to further subdivisions. Notice, nevertheless, the different components of the Eastern Eurasian source populations among Finno-Ugrians:
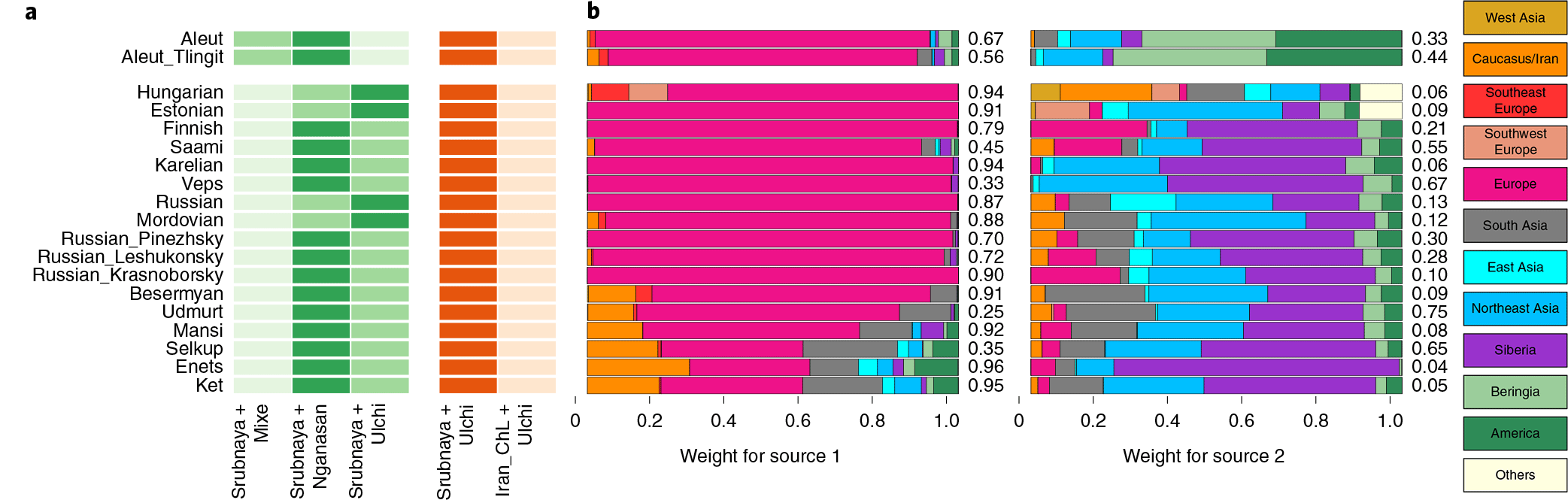
Also remarkable is the lack of comparison of Uralic populations with other neighbouring ones, since the described Uralic-like ancestry of Russians was already known, and is most likely due to the recent acculturation of Uralic-speaking peoples in the cradle of Russians, right before their eastward expansions.
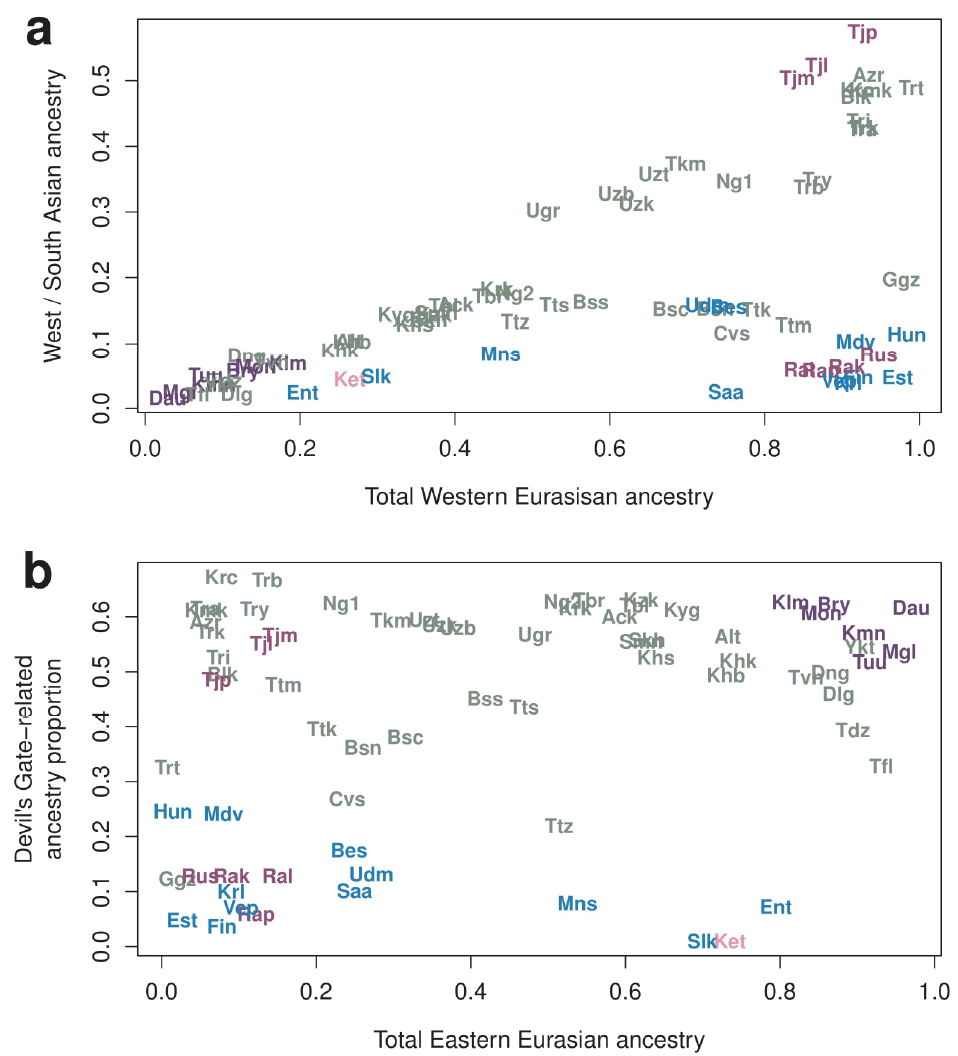
A comparison of Estonians and Finns with Balts, Scandinavians, and Eastern Europeans would have been more informative for the division of the different so-called “Nganasan-like meta-populations”, and to ascertain which one of these ancestral peoples along the ancient WHG:ANE cline could actually be connected (if at all) to the Cis-Urals.
Because, after all, based on linguistics and archaeology, geneticists are not supposed to be looking for populations from the North Asian Arctic region, for “Siberian ancestry”, or for haplogroup N1c – despite previous works by their peers – , but for the Bronze Age Volga-Kama region…
Graphic: using Population Genomics to follow "Steppe ancestry + R1a" as *Indo-Europeans* and "Siberian ancestry + hg. N1c" as *Uralics*. https://t.co/aEeOmEfxzD pic.twitter.com/Y8wm8VN0q9
— Carlos Quiles (@cquilesc) May 17, 2019
See also
Related
- The cradle of Russians, an obvious Finno-Volgaic genetic hotspot
- Common Slavs from the Lower Danube, expanding with haplogroup E1b-V13?
- Aquitanians and Iberians of haplogroup R1b are exactly like Indo-Iranians and Balto-Slavs of haplogroup R1a
- Magyar tribes brought R1a-Z645, I2a-L621, and N1a-L392(xB197) lineages to the Carpathian Basin
- R1a-Z280 and R1a-Z93 shared by ancient Finno-Ugric populations; N1c-Tat expanded with Micro-Altaic
- The complex origin of Samoyedic-speaking populations
- Corded Ware—Uralic (IV): Hg R1a and N in Finno-Ugric and Samoyedic expansions
- Corded Ware—Uralic (III): “Siberian ancestry” and Ugric-Samoyedic expansions
- Corded Ware—Uralic (II): Finno-Permic and the expansion of N-L392/Siberian ancestry
- The traditional multilingualism of Siberian populations
- Corded Ware—Uralic (I): Differences and similarities with Yamna
- Haplogroup R1a and CWC ancestry predominate in Fennic, Ugric, and Samoyedic groups