New open access paper Ancient DNA sheds light on the genetic origins of early Iron Age Philistines, by Feldman et al. Science Advances (2019) 5(7):eaax0061.
Interesting excerpts (modified for clarity, emphasis mine):
Here, we report genome-wide data from human remains excavated at the ancient seaport of Ashkelon, forming a genetic time series encompassing the Bronze to Iron Age transition. We find that all three Ashkelon populations derive most of their ancestry from the local Levantine gene pool. The early Iron Age population was distinct in its high genetic affinity to European-derived populations and in the high variation of that affinity, suggesting that a gene flow from a European-related gene pool entered Ashkelon either at the end of the Bronze Age or at the beginning of the Iron Age. Of the available contemporaneous populations, we model the southern European gene pool as the best proxy for this incoming gene flow. Last, we observe that the excess European affinity of the early Iron Age individuals does not persist in the later Iron Age population, suggesting that it had a limited genetic impact on the long-term population structure of the people in Ashkelon.
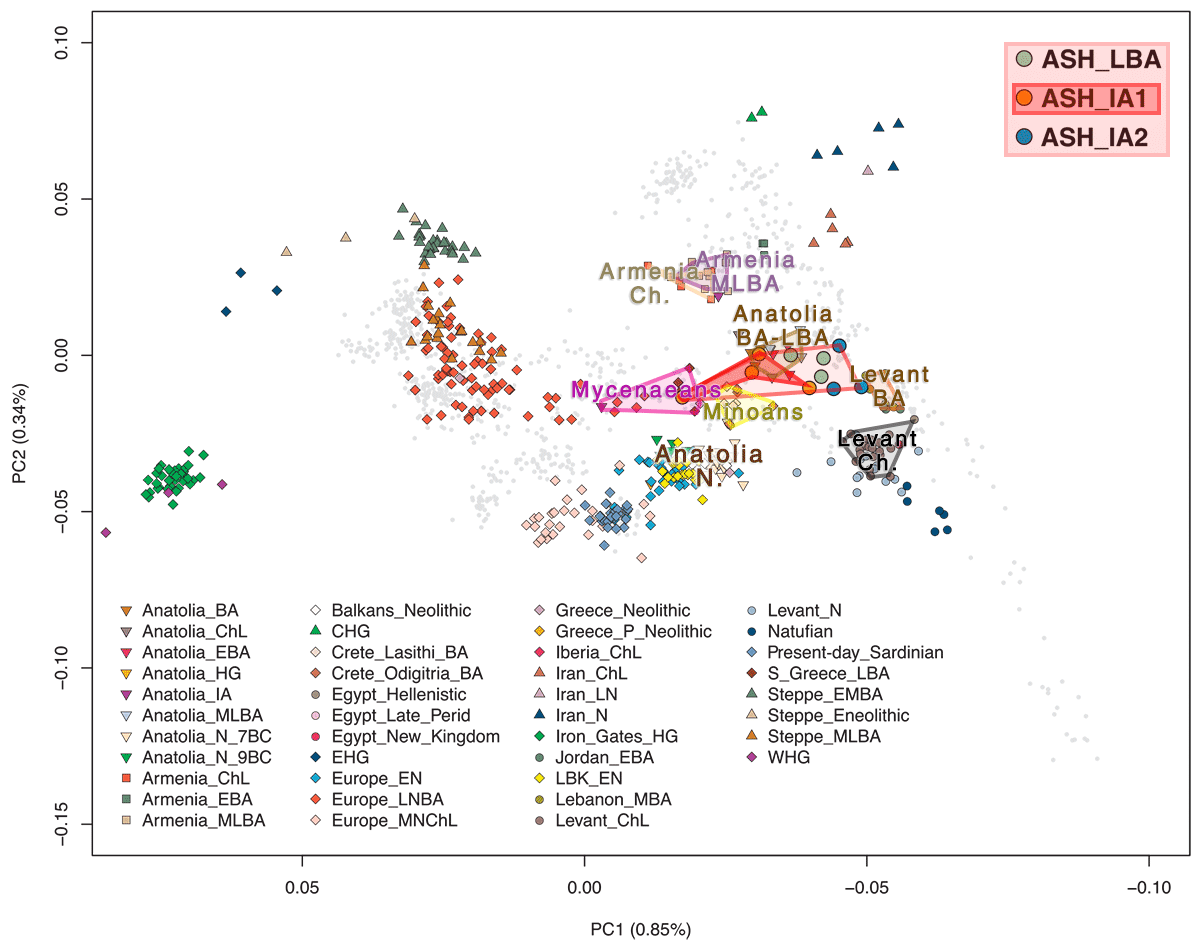
Genetic discontinuity between the Bronze Age and the early Iron Age people of Ashkelon
In comparison to ASH_LBA, the four ASH_IA1 individuals from the following Iron Age I period are, on average, shifted along PC1 toward the European cline and are more spread out along PC1, overlapping with ASH_LBA on one extreme and with the Greek Late Bronze Age “S_Greece_LBA” on the other. Similarly, genetic clustering assigns ASH_IA1 with an average of 14% contribution from a cluster maximized in the Mesolithic European hunter-gatherers labeled “WHG” (shown in blue in Fig. 2B) (15, 22, 26). This component is inferred only in small proportions in earlier Bronze Age Levantine populations (2 to 9%).
In agreement with the PCA and ADMIXTURE results, only European hunter-gatherers (including WHG) and populations sharing a history of genetic admixture with European hunter-gatherers (e.g., as European Neolithic and post-Neolithic populations) produced significantly positive f4-statistics (Z ≥ 3), suggesting that, compared to ASH_LBA, ASH_IA1 has additional European-related ancestry.
We find that the PC1 coordinates positively correlate with the proportion of WHG ancestry modeled in the Ashkelon individuals, suggesting that WHG reasonably tag a European-related ancestral component within the ASH_IA1 individuals.
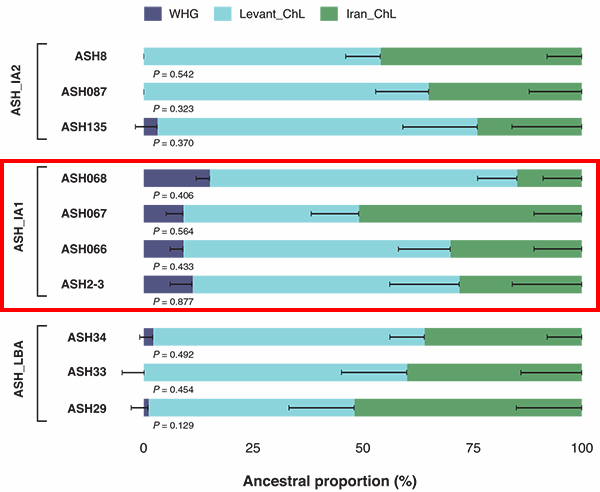
The best supported one (χ2P = 0.675) infers that ASH_IA1 derives around 43% of ancestry from the Greek Bronze Age “Crete_Odigitria_BA” (43.1 ± 19.2%) and the rest from the ASH_LBA population.
(…) only the models including “Sardinian,” “Crete_Odigitria_BA,” or “Iberia_BA” as the candidate population provided a good fit (χ2P = 0.715, 49.3 ± 8.5%; χ2P = 0.972, 38.0 ± 22.0%; and χ2P = 0.964, 25.8 ± 9.3%, respectively). We note that, because of geographical and temporal sampling gaps, populations that potentially contributed the “European-related” admixture in ASH_IA1 could be missing from the dataset.
The transient impact of the “European-related” gene flow on the Ashkelon gene pool
The ASH_IA2 individuals are intermediate along PC1 between the ASH_LBA ones and the earlier Bronze Age Levantines (Jordan_EBA/Lebanon_MBA) in the west Eurasian PCA (Fig. 2A). Notably, despite being chronologically closer to ASH_IA1, the ASH_IA2 individuals position closer, on average, to the earlier Bronze Age individuals.
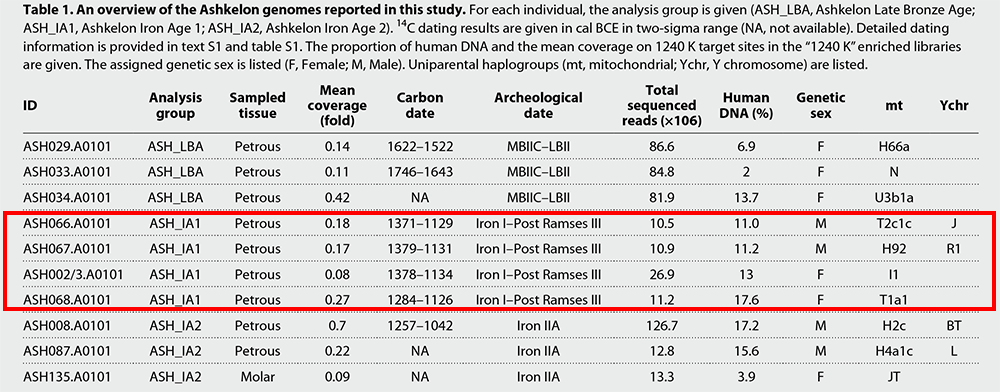
The transient excess of European-related genetic affinity in ASH_IA1 can be explained by two scenarios. The early Iron Age European-related genetic component could have been diluted by either the local Ashkelon population to the undetectable level at the time of the later Iron Age individuals or by a gene flow from a population outside of Ashkelon introduced during the final stages of the early Iron Age or the beginning of the later Iron Age.
By modeling ASH_IA2 as a mixture of ASH_IA1 and earlier Bronze Age Levantines/Late Period Egyptian, we infer a range of 7 to 38% of contribution from ASH_IA1, although no contribution cannot be rejected because of the limited resolution to differentiate between Bronze Age and early Iron Age ancestries in this model.
Hg. R1b-M269 and the Aegean
I already predicted this relationship of Philistines and Aegeans (Greeks in particular) months ago, based on linguistics, archaeology, and phylogeography, although it was (and still is) yet unclear if these paternal lineages might have come from other nearby populations which might be descended from Common Anatolians instead, given the known intense contacts between Helladic and West Anatolian groups.
The deduction process for the Greek connection was quite simple:
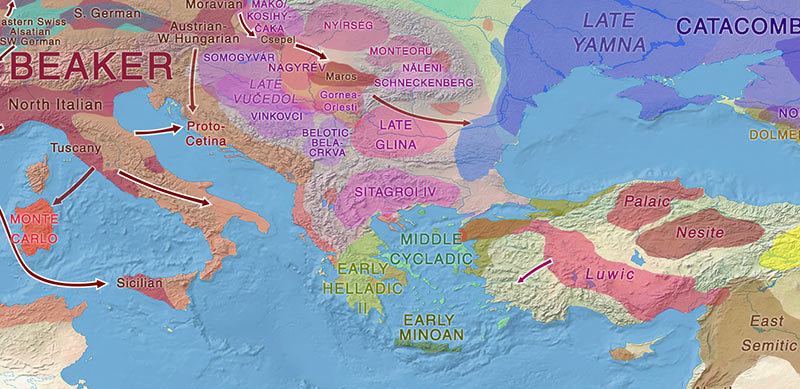
Palaeo-Balkan populations
We know that R1b-Z2103 expanded with Yamna, including West Yamna settlers: they appear in Vučedol, which means they formed part of the earliest expansion waves of Yamna settlers into the Carpathian Basin, and they also appear scattered among Bell Beakers (apart from dominating East Yamna and Afanasevo), which suggests that they were possibly one of the most successful lineages during the late Repin/early Yamna expansion.
The “Steppe ancestry” associated with I2a-L699 samples among Balkan BA peoples may have also been associated with recent Bronze Age expansions, and this haplogroup’s presence among modern Balkan peoples may also suggest that it expanded with Palaeo-Balkan languages. Nevertheless, we don’t know which specific lineages and “Steppe ancestry” they represent, sadly.
These samples may well be related to remnants of previous Balkan populations like Cernavodă or Ezero, because there has been no peer-reviewed attempt at distinguishing Khvalynsk-/Novodanilovka- from Sredni Stog- from Yamnaya-related populations (see here), and some groups that are associated with this ancestry, like Corded Ware, are known to be culturally distinct from Yamna.
In any case, Proto-Greeks from the southern Balkans (say, Sitagroi IV and related groups) are probably going to show, based on Palaeo-Balkan substrate and Pre-Greek substrate and on the available Mycenaean samples, a process of decreasing proportion of R1b-Z2103 lineages relative to local ones, and a relatively similar cline of Yamna:EEF ancestry from northern to southern areas, at least in the periods closest to the Yamna expansion.
NOTE. The finding of “archaic” R1b-L389 (R1b-V1636) and R1a-M198 subclades among modern Greeks and the likely Neolithic origin of these paternal lineages around the Caucasus suggest that their presence in Greece may be from any of the more recent migrations that have happened between Anatolia and the Balkans, especially during the Common Era, rather than Indo-Anatolian migrations; probably very very recently.
Minoans and haplogroup J
In the Aegean, it is already evident that the population changed language partly through cultural diffusion, probably through elite domination of Proto-Greek speakers. Whether that happened before the invasion into the Greek Peninsula or after it is unclear, as we discussed recently, because we only have one reported Y-chromosome haplogroup among Mycenaeans, and it is J (probably continuing earlier lineages).
Now we have more samples from the so-called Emporion 2 cluster in Olalde et al. (2019), which shows Mycenaean-like eastern Mediterranean ancestry and 3 (out of 3) samples of haplogroup J, which – given the origin of the colony in Phocea – may be interpreted as the prevalence of West Anatolian-like ancestry and lineages in the eastern part of the Aegean (and possibly thus south Peloponnese), in line with the modern situation.
NOTE. It does not seem likely that those R or R1b-L23 samples from the Emporion 1 cluster are R1b-Z2103, based on their West European-like ancestry, although they still may be, because – as we know – ancestry (unlike haplogroup) changes too easily to interpret it as an ancestral ethnolinguistic marker.
Greeks and haplogroup R1b-M269
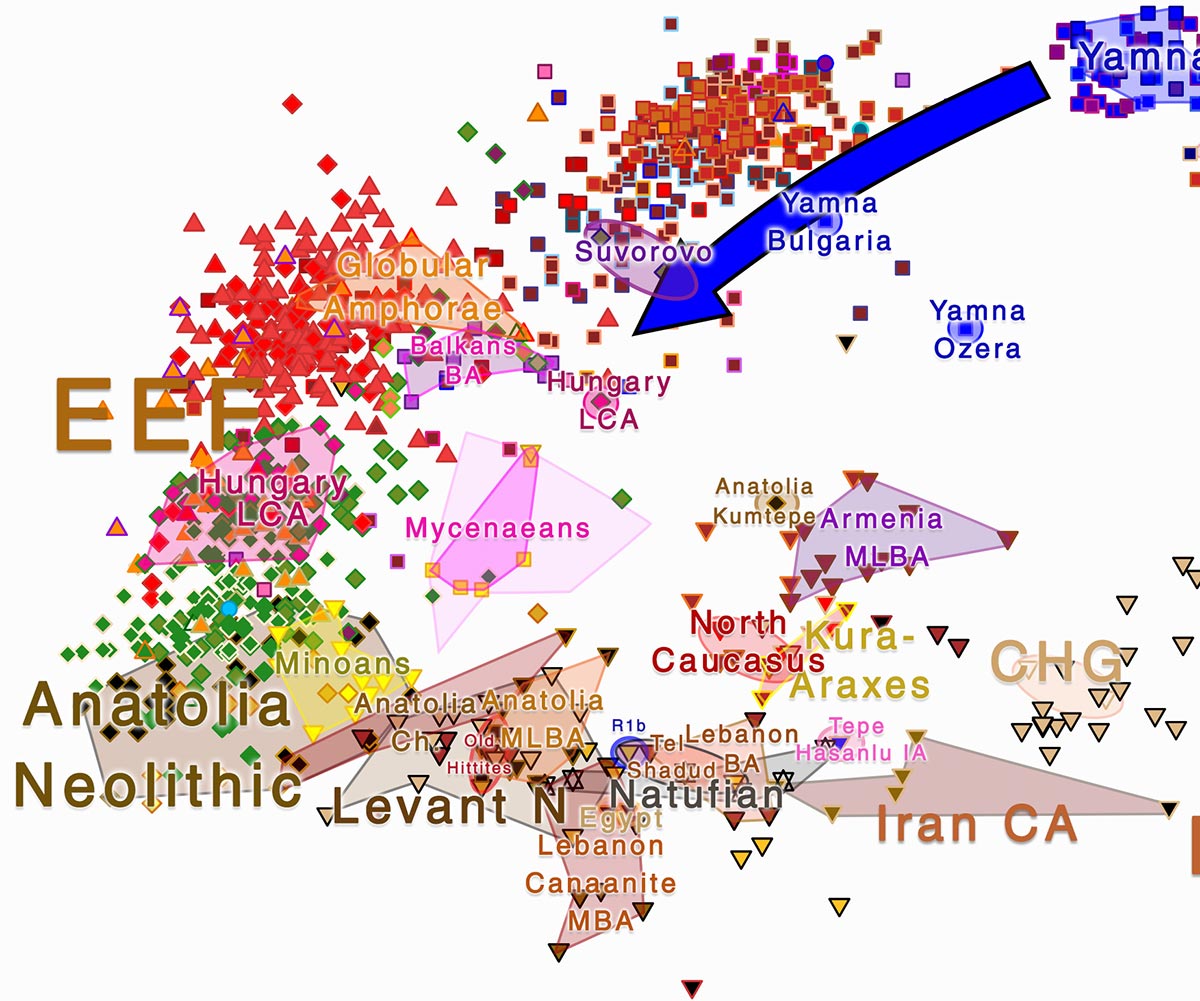
Therefore, while the presence of R1b-Z2103 among ancient Balkan peoples connected to the Yamna expansion is clear, one might ask if R1b-Z2103 really spread up to the Peloponnese by the time of the Mycenaean Civilization. That has only one indirect answer, and it’s most likely yes.
We already had some R1b-Z2103 among Thracians and around the Armenoid homeland, which offers another clue at the migration of these lineages from the Balkans. The distribution of different “archaic” R1b-Z2103 subclades among modern Balkan populations and around the Aegean offered more support to this conclusion.
But now we have two interesting ancient populations that bear witness to the likely intrusion of R1b-M269 with Proto-Greeks:
An Ancient Greek of hg. R1b
A single ancient sample supports the increase in R1b-Z2103 among Greeks during the “Dorian” invasions that triggered the Dark Ages and the phenomenon of the Aegean Sea Peoples. It comes from a Greek lab study, showing R1b1b (i.e. R1b-P297 in the old nomenclature) as the only Y-chromosome haplogroup obtained from the sampling of the Gulf of Amurakia ca. 470-30 BC, i.e. before the Roman foundation of Nikopolis, hence from people likely from Anaktorion in Ancient Acarnania, of Corinthian origin.
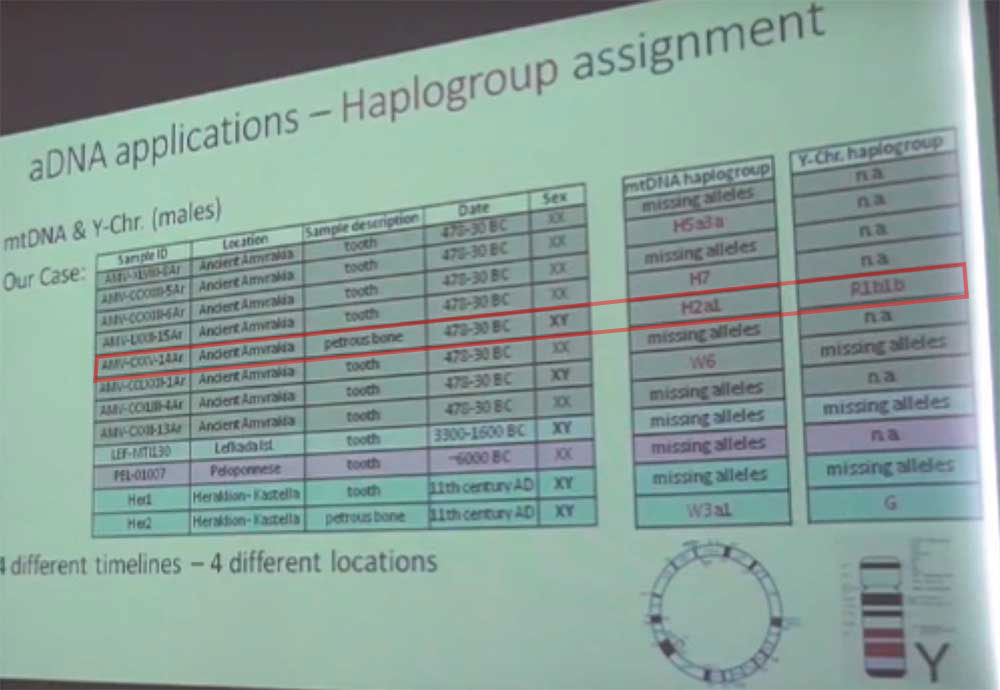
Even with the few data available – and with the caution necessary for this kind of studies from non-established labs, which may be subject to many different kinds of errors – one could argue that the western Greek areas, which received different waves of migrants from the north and shows a higher distribution of R1b-Z2103 in modern times, was probably more heavily admixed with R1b-Z2103 than southern and eastern areas, which were always dominated by Greek-speaking populations more heavily admixed with locals.
The Dorian invasion and the Greek Dark Ages may thus account for a renewed influx of R1b-Z2103 lineages accompanying the dialects that would eventually help form the Hellenic Koiné. In a sense, it is only natural that demographically stronger populations around the Bronze Age Aegean would suffer a limited (male) population replacement with the succeeding invasions, starting with a higher genetic impact in the north-west and diminishing as they progressed to the south and the east, coupled with stepped admixture events with local populations.
This would be therefore the late equivalent of what happened at the end of the 3rd millennium BC, with Mycenaeans and their genetic continuity with Minoans.
Sea peoples of hg. R1b-M269
Thanks to Wang et al. (2018) supplementary materials we knew that one of the two Levantine LBA II samples from Tel Shadud (final 13th–early 11th c. BC) published in van den Brink (2017) was of hg. R1b-M269 – in fact, the one interpreted as a Canaanite official residing at this site and emulating selected funerary aspects of Egyptian mortuary culture.
Both analyzed samples, this elite individual and a commoner of hg. J buried nearby, were genetically similar and indistinguishable from local populations, though:
Principal Components Analysis of L112 and L126 was carried out within the framework described in Lazaridis et al. (2016). This analysis showed that the two individuals cluster genetically, with similar estimated proportions of ancestry from diverse West Eurasian ancestral sources. These results are consistent with the hypothesis that they derive from the same population, or alternatively that they derive from two quite closely related populations.
We know that ancestry changes easily within a few generations, so there was not much information to go on, except for the fact that – being R1b-M269 – this individual could trace his paternal ancestor at some point to Proto-Indo-Europeans.
One might think that, because many haplogroups in this spreadsheet were wrong, this is also wrong; nevertheless, many haplogroups are correctly identified by Yleaf, and finding R1b-M269 in the Levant after the expansion of Sea Peoples could not be that surprising, because they were most likely related to populations of the Aegean Sea. Any other related hg. R1b (R1b-M73, R1b-V88, even R1b-V1636) wouldn’t fit as well as R1b-M269.

However, the early expansion of Proto-Indo-Aryans into the Middle East, as well as the later expansion of Armenians from the Balkans through Anatolia and of West Iranians from the east may have all potentially been related to this sample. But still, the previous linguistic and archaeological theories concerning the Philistines and the expansion of Sea Peoples in the Levant made this sample a likely (originally) Greek “Dorian” lineage, rather than the other (increasingly speculative) alternatives.
In any case, it was obvious to anyone – that is, to anyone with a minimum knowledge of how population genomics works – that just the two samples from van den Brink (2017) couldn’t be used to get to any conclusions about the ancestral origin of these individuals (or their differences) beyond Levantine peoples, because their ancestry was essentially (i.e. statistically) the same as the other few available ancient samples from nearby regions and similar periods.
If anything, the PCA suggested an origin of the R1b sample closer to Aegean populations relative to the J individual (see PCA above), and this should have been supported also by amateur models, without any possible confirmation (as with the ASH_IA2 cluster in this paper). However, if you have followed online discussions of Tel Shadud R1b-M269 sample since it was mentioned first on Eupedia months ago – including another wave of misguided speculation based on the ancestry of both individuals triggered by a discussion on this blog -, you have once more proof of how misleading ancestry analyses can be in the wrong hands.
NOTE. This is the Nth proof (and that only in 2019) of how it’s best to just avoid amateur analyses and interpretations altogether, as I did in the recent publication of the books. All those who didn’t take into account whatever was commented about the ancestry of these samples haven’t lost a single bit of relevant information on Levantine peoples, and have had more time for useful reads, compared to those dedicated to endless void speculation, once again gone awfully wrong, as does everything related to cocky ancient DNA crackpottery ;-)
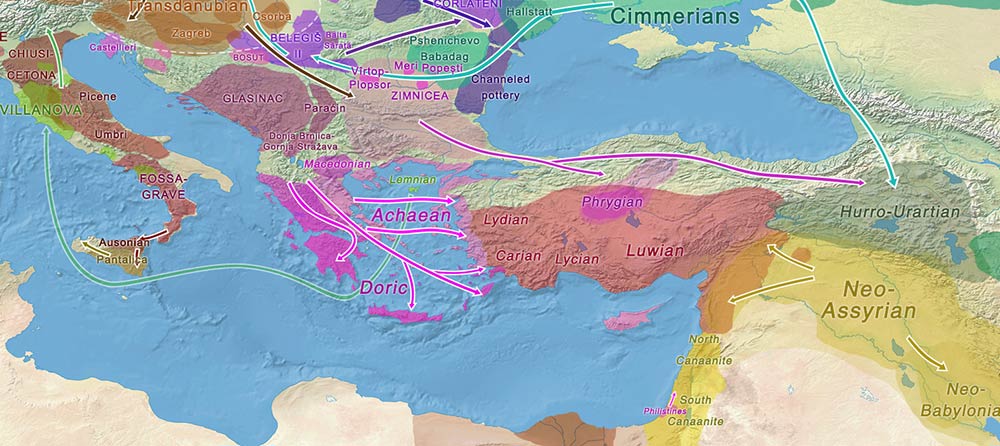
Admittedly, though, even accepting the evident Mediterranean origin of this lineage, one could have argued that this sample may have been of R1b-L151 subclade, if one were inclined to support the theory that Italic peoples were behind Sea Peoples expanding east – and consequently that the ancestors of Etruscans had migrated eastward into the Aegean (e.g. into Lemnos), so that it could be asserted that Tyrsenian might have been a remnant language of an ancient population of northern Italy.
Philistines
Fortunately, some of the samples recovered in Feldman et al. (2019) that could be analyzed (those of the cluster ASH_IA1) offer a very specific time frame where European ancestry appeared (ca. 1250 BC) before it subsequently became fully diluted (as seen in cluster ASH_IA2) among the prevalent Levantine ancestry of the area.
Also fortunately, this precise cluster shows another R1b-M269 sample, likely R1b-Z2103 (because it is probably xL151), and this sample together with others from the same cluster prove that the ancestry related to the original southern European incomers was:
- Recent, related thus to LBA population movements, as expected; and
- More closely related to coeval Aegeans, including Mycenaeans with Steppe-related ancestry.
NOTE. I say “fortunately” because, as you can imagine if you have dealt with amateurish discussions long enough, without this cluster with evident Aegean ancestry and the R1b-M269 (Z2103) sample precisely associated to it, some would enter again in endless comment loops created by ancestry magicians, showing how Aegean peoples were not behind Sea Peoples, or not behind Philistines, or not behind the R1b-M269 among Philistines, depending on their specific agendas.
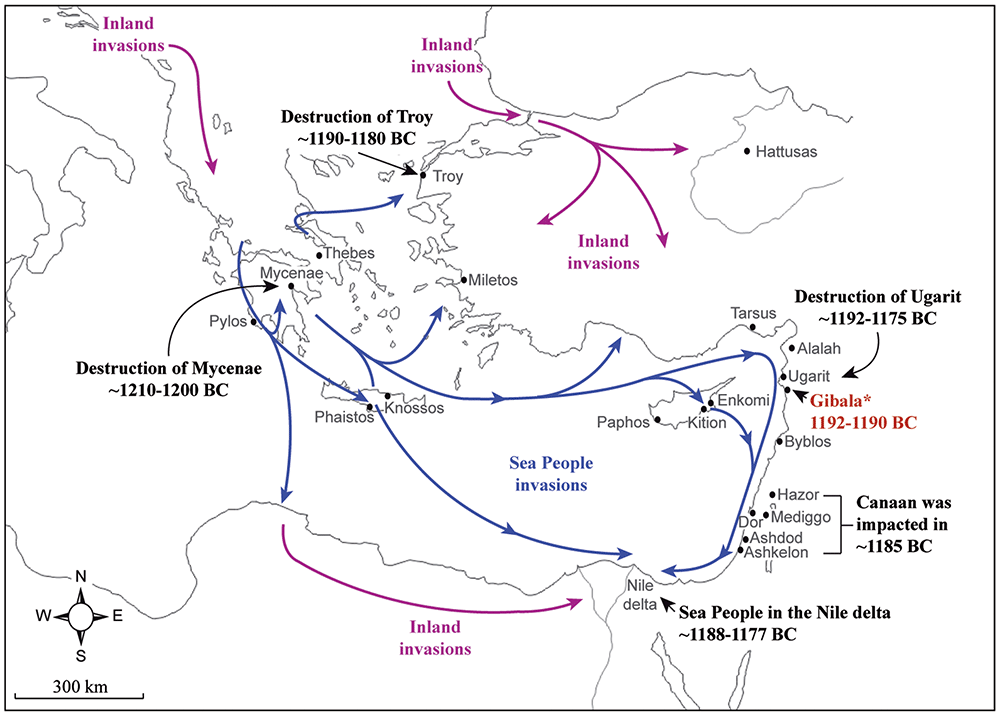
The results of the paper don’t solve the question of the exact origin of all Sea Peoples (not even that of Philistines), but it is quite clear that most of those forming this seafaring confederation must have come from sites around the Aegean Sea. This supports thus the traditional origin attributed to them, including a hint at the likely expansion of Eastern Mediterranean ancestry and lineages into the Italian Peninsula precisely from the Aegean, as some oral communications have already disclosed.
As an indirect conclusion from the findings in this paper, then, we can now more confidently support that Tyrsenian speakers most likely expanded into the Appenines and the Alps originally from a Tyrsenian-speaking LBA population from Lemnos, due to the social unrest in the whole Aegean region, and might have become heavily admixed with local Italic peoples quite quickly, as it happened with Philistines, resulting in yet another case of language expansion through (the simplistically called) elite domination.
Conclusion
Even more interesting than these specific findings, this paper confirms yet another hypothesis based on phylogeography, and proves once again two important starting points for ancient DNA interpretation that I have discussed extensively in this blog:
- The rare R1b-M269 Y-chromosome lineage of Tel Shadud offered ipso facto the most relevant clue about the ancestral geographical origin of this Canaanite elite male’s paternal family, most likely from the north-west based on ancient phylogeography, which indirectly – in combination with linguistics and archaeology – supported the ancestral ethnolinguistic identification of Philistines with the Aegean and thus with (a population closest to) Ancient Greeks.
- Ancestry analyses are often fully unreliable when assessing population movements, especially when few samples from incomplete temporal-geographical transects are assessed in isolation, because – unlike paternal (and maternal) haplogroups – ancestry might change fully within a few generations, depending on the particular anthropological setting. Their investigation is thus bound by many limitations – of design, statistical, and anthropological (i.e. archaeological and linguistic) – which are quite often not taken into account.
These cornerstones of ancient DNA interpretation have been already demonstrated to be valid not only for Levantine populations, as in this case, but also for Balkan peoples, for Bell Beakers, for steppe populations (like Khvalynsk, Sredni Stog, Yamna, Corded Ware), for Basques, for Balto-Slavs, for Ugrians and Samoyeds, and for many other prehistoric peoples.
I rest my case.
Related
- Iberia: East Bell Beakers spread Indo-European languages; Celts expanded later
- Genetic origins of Minoans and Mycenaeans and their continuity into modern Greeks
- On the origin of R1a and R1b subclades in Greece
- Ancient Sardinia hints at Mesolithic spread of R1b-V88, and Western EEF-related expansion of Vasconic
- Arrival of steppe ancestry with R1b-P312 in the Mediterranean: Balearic Islands, Sicily, and Iron Age Sardinia
- Y-DNA relevant in the postgenomic era, mtDNA study of Iron Age Italic population, and reconstructing the genetic history of Italians
- Modern Sardinians show elevated Neolithic farmer ancestry shared with Basques
- Ancient Phoenician mtDNA from Sardinia, Lebanon reflects settlement, genetic diversity, and female mobility
- Haplogroup J spread in the Mediterranean due to Phoenician and Greek colonizations
- Migration vs. Acculturation models for Aegean Neolithic in Genetics — still depending strongly on Archaeology
- Aquitanians and Iberians of haplogroup R1b are exactly like Indo-Iranians and Balto-Slavs of haplogroup R1a