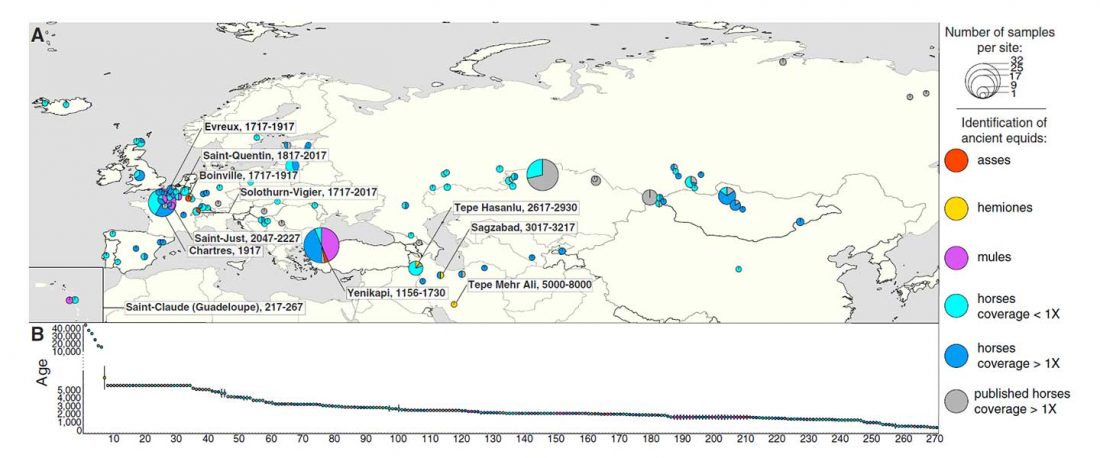
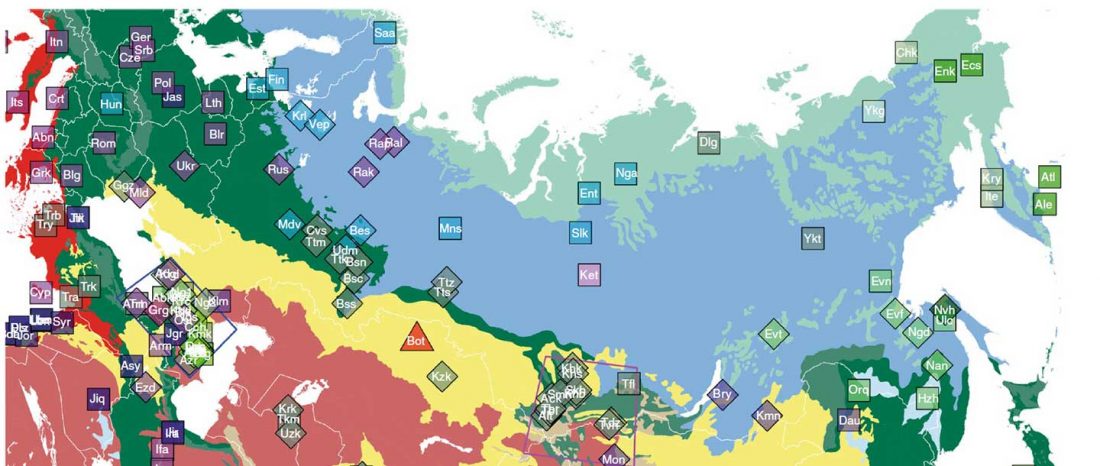
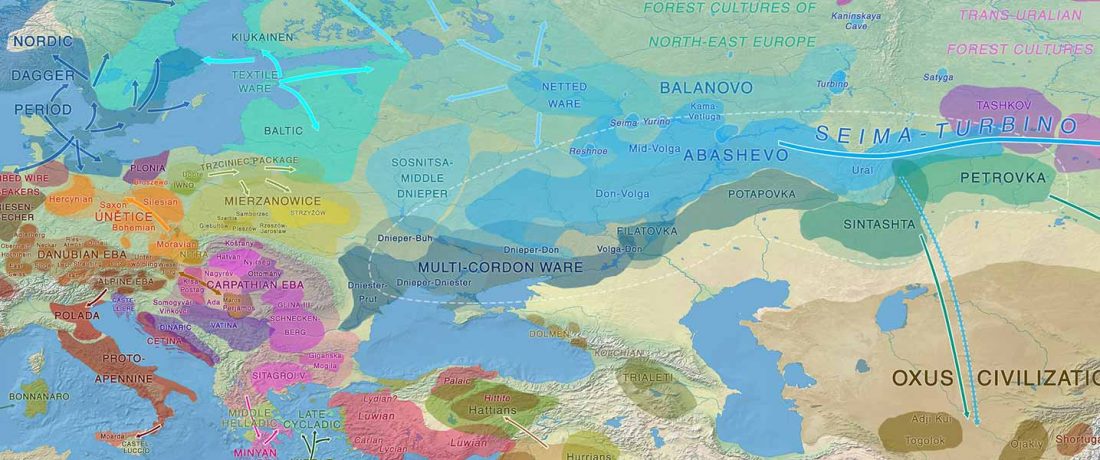
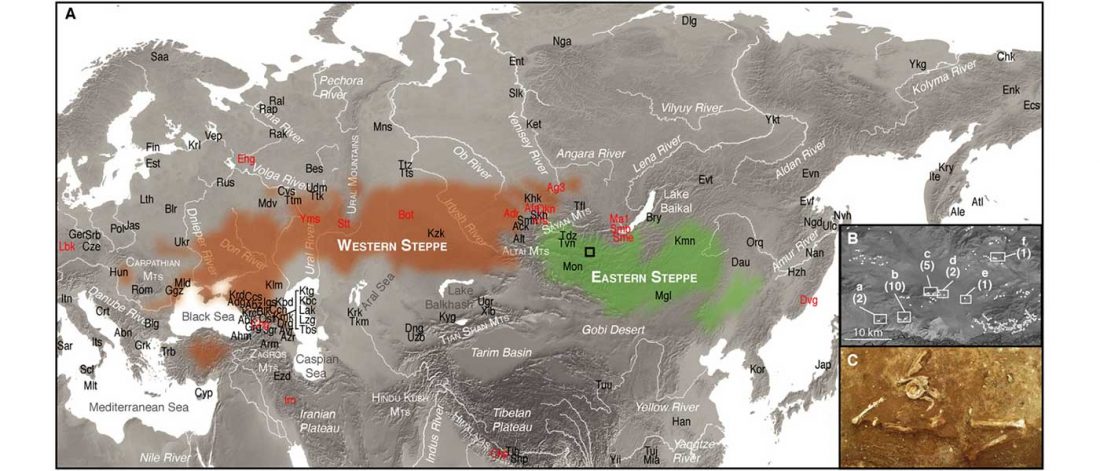
This post is part of a draft on palaeolinguistics and the Proto-Uralic homeland. See below for the color code of protoforms.
10. Metallurgy
PU (Saa., Fi., Md., Ma., P, Ms.?, Kh., Smy.?) *wäśkä (*waśki?) ‘copper; ore, brass’ (UEW Nº 1123; Kallio 2006: 6). Irregular cognates suggest it might have been borrowed during the split-up of Proto-Uralic (cf. Aikio 2015: 42). However, compare potentially regular cognates from *wäskä in PFi. *vaski ‘ore, copper, bronze; brass’ (Kallio 2012: 167; Zhivlov 2014: 115), PSaa. *weśkä ‘copper; brass’, Md. Kazhlodka viśkä ‘chain’ (Häkkinen 2012: 18), and possibly Hu. … Read the rest “Proto-Uralic Homeland (VI): Mythology & Metallurgy”