Open access Siberian genetic diversity reveals complex origins of the Samoyedic-speaking populations, by Karafet et al. Am J Hum Biol (2018) e23194.
Interesting excerpts (emphasis mine):
Siberian groups
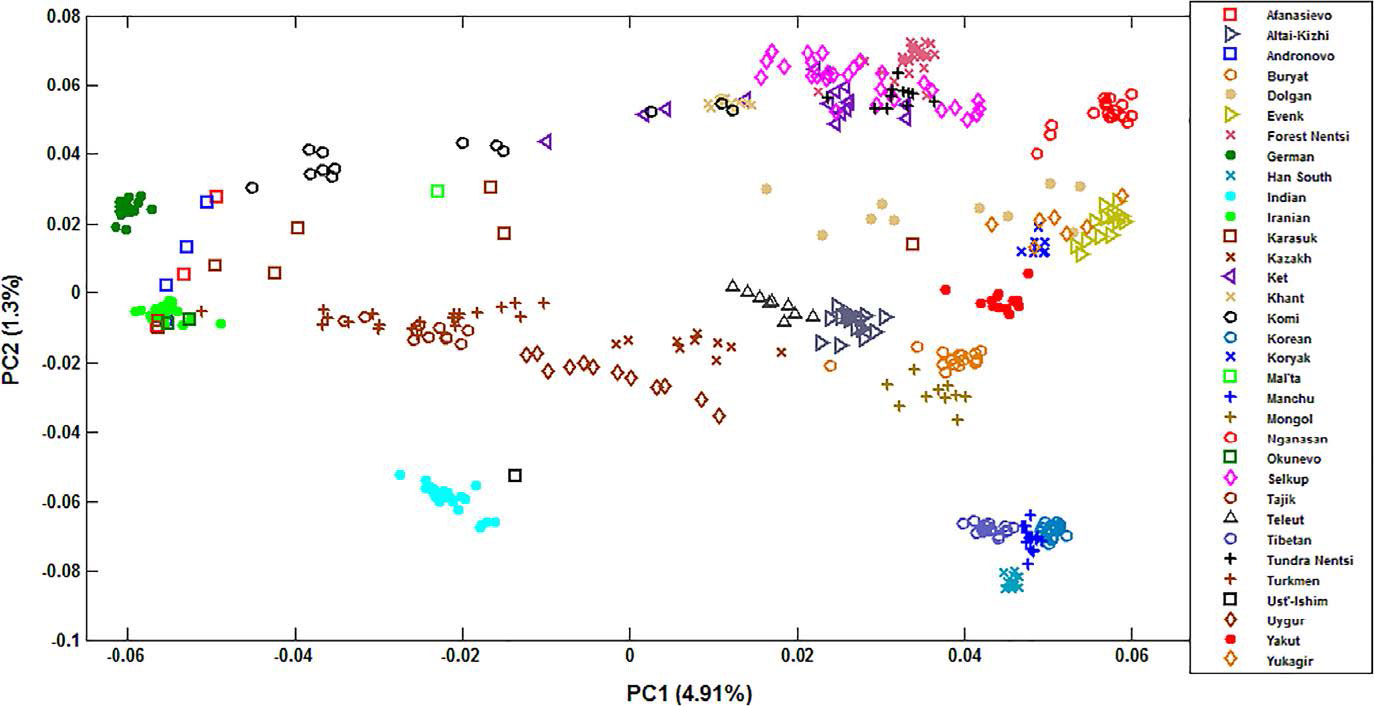
Consistent with their origin, Mongolic-speaking Buryats demonstrate genetic similarity with Mongols, and Turkic-speaking Altai-Kizhi and Teleuts are drawn close to CAS groups. The Tungusic-speaking Evenks collected in central and eastern Siberia cluster together and overlap with Yukagirs. Dolgans are widely scattered in the plot, justifying their recent origin from one Evenk clan, Yakuts, and Russian peasants in the 18th century (Popov, 1964). Uralic-speaking populations comprise a very wide cluster with Komi drawn to Europe, and Khants showing a closer affinity with Selkups, Tundra and Forest Nentsi. Yenisey-speaking Kets are intermingled with Selkups. Interestingly, Samoyedic-speaking Nganasans from the Taymyr Peninsula form a separate tight cluster closer to Evenks, Yukagirs, and Koryaks.
ADMIXTURE and the “Siberian component”
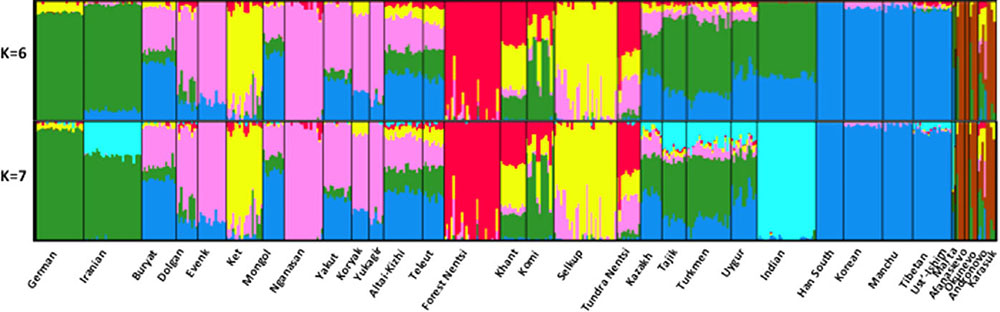
Among Siberians, the Komi are primarily Europeans, while Nganasans, Evenks, Yukagirs, and Koryaks are nearly 100% East Asians. At K = 4 finer scale subcontinental structure can be distinguished with the emergence of a “Siberian” component. This component is highly pronounced in the Nganasans. Outside Siberia, this component is present in Germany and in CAS at low frequency. Within ancient cultures, this component has the highest frequency in three BA Karasuk samples. It is also found in Mal’ta, ENE Afanasievo and BA Andronovo, but not in Ust’-Ishim and BA Okunevo. At K = 5, the “Siberian” component is roughly subdivided into two components with different geographic distributions. The “Nganasan” component is frequent in nearly all Siberian populations, except the Komi, Kets and Selkups. The newly derived “Selkup-Ket” component is found at high frequencies in western Siberian populations. It is observed in BA Karasuk and in Mal’ta. At K = 6, the western Siberian “Nentsi-Khant” ancestry component was developed in Forest and Tundra Nentsi, Khants. This component is also present at low levels in EUR, CAS, Tibet, and southern Siberia.
Identity-by-descent
The Dolgans share more segments with the Nganasans than within themselves (54.13 vs 41.72, Mann-Whitney test, P = .000000000001562546). The result is not surprising as the demographic data showed that the Nganasans were subjected to intense assimilation by the Dolgans in the second half of the 20th century (Goltsova, Osipova, Zhadanov, & Villems, 2005). Tundra Nentsi share more IBD with Forest Nentsi than within themselves (83.96 vs 50.3, P = .000055) possibly due to the common origin and long-term gene flow. The Ket and Selkup populations allocate significantly more IBD blocks between populations than with individuals from their own population (121.2 cM vs 85.9 cM for Kets, P = .000008, and 121.2 cM vs 114.9 cM for Selkups, P = .043).
Haplogroup N in Siberia
Although Siberia exhibits 42 haplogroups, the vast majority of Siberian Y-chromosomes belong only to 4 of the 18 major clades (N = 46.2%; C = 20.9%; Q = 14.4%; and R = 15.2%). The Y-chromosome haplogroup N is widely spread across Siberia and Eastern Europe (Ilumae et al., 2016; Karafet et al., 2002; Wong et al., 2016) and reaches its maximum frequency among Siberian populations such as Nganasans (94.1%) and Yakuts (91.9%). Within Siberia, two sister subclades N-P43 and N-L708 show different geographic distributions. N-P43 and derived haplogroups N-P63 and N- P362 (phylogenetically identical to N-B478* and N-B170, respectively) (Ilumae et al., 2016) are extremely rare in other major geographic regions. Likely originating in western Siberia, they are limited almost entirely to northwest Siberia, the Volga- Uralic regions, and the Taymyr Peninsula (ie, do not extend to eastern Siberia). Conversely, clade N-L708 is frequent in all Siberian populations except the Kets and Selkups, reaching its highest frequency in the Yakuts (91.9%).
Surprisingly, not a single sign of the proposed reindeer pastoralist horde led by Nganasans into north-eastern Europe. This is strange because “Siberian” migrants hypothetically imposed their language over Indo-Europeans quite recently, apparently after the Iron Age…
Interesting comparisons among Siberian groups, though.
Related
- Corded Ware—Uralic (IV): Hg R1a and N in Finno-Ugric and Samoyedic expansions
- Corded Ware—Uralic (III): “Siberian ancestry” and Ugric-Samoyedic expansions
- Corded Ware—Uralic (II): Finno-Permic and the expansion of N-L392/Siberian ancestry
- The traditional multilingualism of Siberian populations
- Iron Age bottleneck of the Proto-Fennic population in Estonia
- Y-DNA haplogroups of Tuvinian tribes show little effect of the Mongol expansion
- Corded Ware—Uralic (I): Differences and similarities with Yamna
- Haplogroup R1a and CWC ancestry predominate in Fennic, Ugric, and Samoyedic groups
- On the origin and spread of haplogroup R1a-Z645 from eastern Europe
- Oldest N1c1a1a-L392 samples and Siberian ancestry in Bronze Age Fennoscandia