Open access paper Bronze Age population dynamics and the rise of dairy pastoralism on the eastern Eurasian steppe, by Jeong et al. PNAS (2018).
Interesting excerpts (emphasis mine):
To understand the population history and context of dairy pastoralism in the eastern Eurasian steppe, we applied genomic and proteomic analyses to individuals buried in Late Bronze Age (LBA) burial mounds associated with the Deer Stone-Khirigsuur Complex (DSKC) in northern Mongolia. To date, DSKC sites contain the clearest and most direct evidence for animal pastoralism in the Eastern steppe before ca. 1200 BCE.
Most LBA Khövsgöls are projected on top of modern Tuvinians or Altaians, who reside in neighboring regions. In comparison with other ancient individuals, they are also close to but slightly displaced from temporally earlier Neolithic and Early Bronze Age (EBA) populations from the Shamanka II cemetry (Shamanka_EN and Shamanka_EBA, respectively) from the Lake Baikal region. However, when Native Americans are added to PC calculation, we observe that LBA Khövsgöls are displaced from modern neighbors toward Native Americans along PC2, occupying a space not overlapping with any contemporary population. Such an upward shift on PC2 is also observed in the ancient Baikal populations from the Neolithic to EBA and in the Bronze Age individuals from the Altai associated with Okunevo and Karasuk cultures.
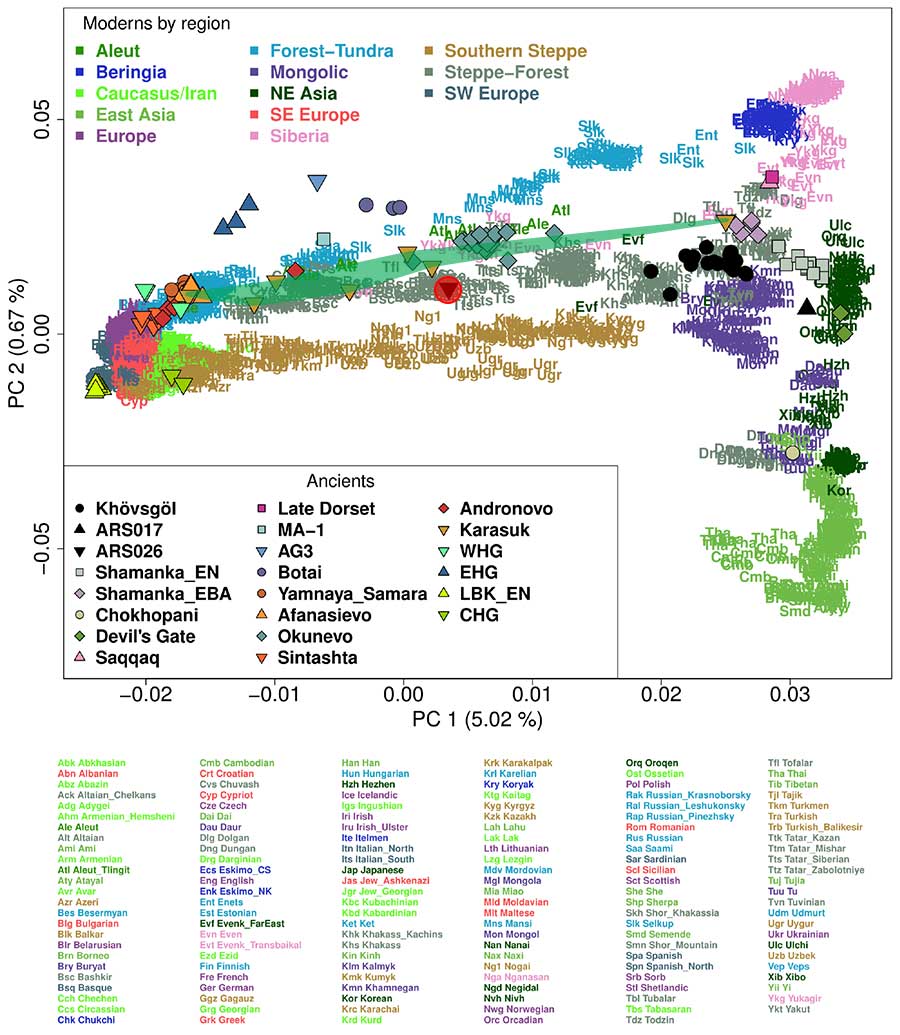
(…) two individuals fall on the PC space markedly separated from the others: ARS017 is placed close to ancient and modern northeast Asians, such as early Neolithic individuals from the Devil’s Gate archaeological site (22) and present-day Nivhs from the Russian far east, while ARS026 falls midway between the main cluster and western Eurasians.
Upper Paleolithic Siberians from nearby Afontova Gora and Mal’ta archaeological sites (AG3 and MA-1, respectively) (25, 26) have the highest extra affinity with the main cluster compared with other groups, including the eastern outlier ARS017, the early Neolithic Shamanka_EN, and present-day Nganasans and Tuvinians (Z > 6.7 SE for AG3). Main cluster Khövsgöl individuals mostly belong to Siberian mitochondrial (A, B, C, D, and G) and Y (all Q1a but one N1c1a) haplogroups.
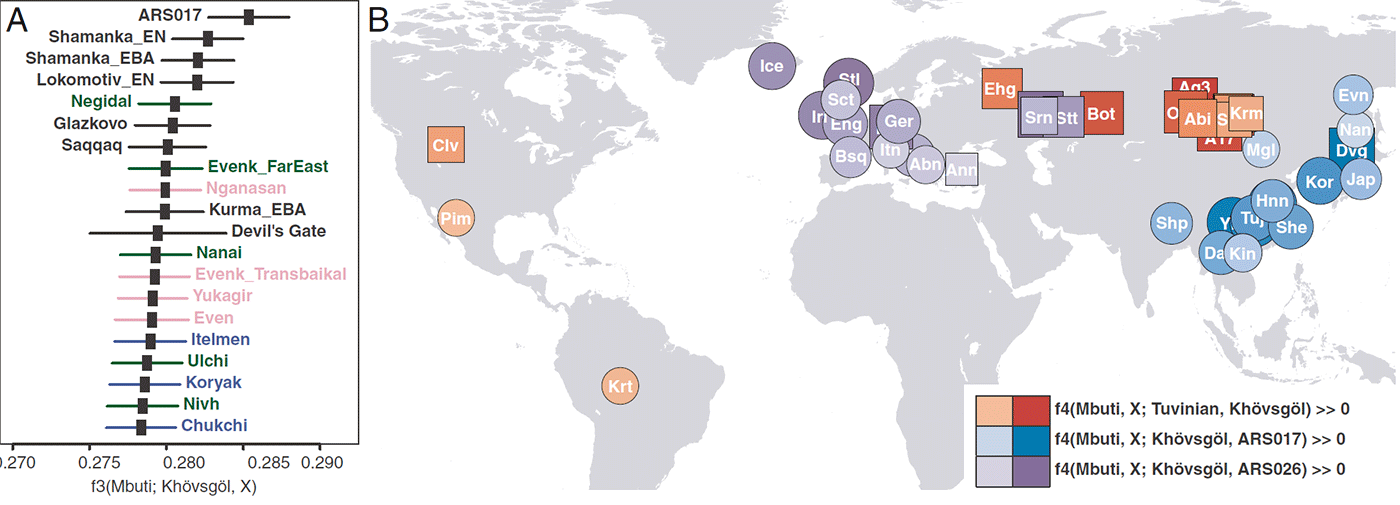
Previous studies show a close genetic relationship between WSH populations and ANE ancestry, as Yamnaya and Afanasievo are modeled as a roughly equal mixture of early Holocene Iranian/ Caucasus ancestry (IRC) and Mesolithic Eastern European hunter-gatherers, the latter of which derive a large fraction of their ancestry from ANE. It is therefore important to pinpoint the source of ANE-related ancestry in the Khövsgöl gene pool: that is, whether it derives from a pre-Bronze Age ANE population (such as the one represented by AG3) or from a Bronze Age WSH population that has both ANE and IRC ancestry.
The amount of WSH contribution remains small (e.g., 6.4 ± 1.0% from Sintashta). Assuming that the early Neolithic populations of the Khövsgöl region resembled those of the nearby Baikal region, we conclude that the Khövsgöl main cluster obtained ∼11% of their ancestry from an ANE source during the Neolithic period and a much smaller contribution of WSH ancestry (4–7%) beginning in the early Bronze Age.
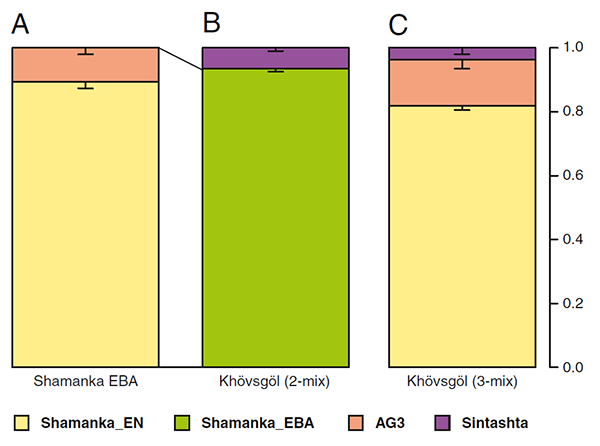
Apparently, then, the first individual with substantial WSH ancestry in the Khövsgöl population (ARS026, of haplogroup R1a-Z2123), directly dated to 1130–900 BC, is consistent with the first appearance of admixed forest-steppe-related populations like Karasuk (ca. 1200-800 BC) in the Altai. Interestingly, haplogroup N1a1a-M178 pops up (with mtDNA U5a2d1) among the earlier Khövsgöl samples.
I will repeat what I wrote recently here: Samoyedic arrived in the Altai with Karasuk and hg R1a-Z645 + Steppe_MLBA-like ancestry, admixed with Altai populations, clustering thus within an Ancient Altai cline. Only later did N1a1a subclades infiltrate Samoyedic (and Ugric) populations, bringing them closer to their modern Palaeo-Siberian cline. The shared mtDNA may support an ancestral EHG-“Siberian” cline, or else a more recent Afanasevo-related origin.
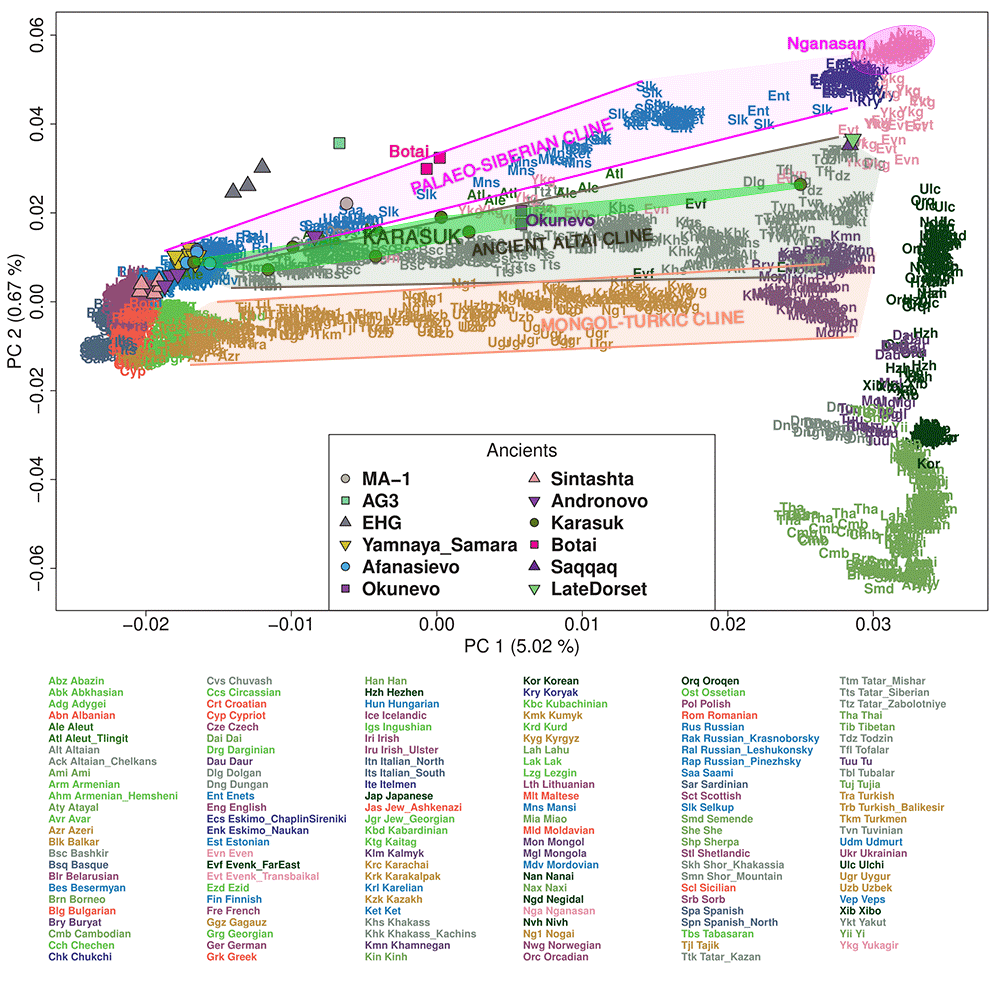
Also interesting, Q1a2 subclades and ANE ancestry making its appearance everywhere among ancestral Eurasian peoples, as Chetan recently pointed out.
Related
- Ancient nomadic tribes of the Mongolian steppe dominated by a single paternal lineage
- Y-DNA haplogroups of Tuvinian tribes show little effect of the Mongol expansion
- Corded Ware—Uralic (II): Finno-Permic and the expansion of N-L392/Siberian ancestry
- Corded Ware—Uralic (III): “Siberian ancestry” and Ugric-Samoyedic expansions
- Corded Ware—Uralic (IV): Haplogroups R1a and N in Finno-Ugric and Samoyedic
- Haplogroup R1a and CWC ancestry predominate in Fennic, Ugric, and Samoyedic groups
- The Iron Age expansion of Southern Siberian groups and ancestry with Scythians
- Evolution of Steppe, Neolithic, and Siberian ancestry in Eurasia (ISBA 8, 19th Sep)
- Mitogenomes from Avar nomadic elite show Inner Asian origin
- Oldest N1c1a1a-L392 samples and Siberian ancestry in Bronze Age Fennoscandia
- Genetic prehistory of the Baltic Sea region and Y-DNA: Corded Ware and R1a-Z645, Bronze Age and N1c
- More evidence on the recent arrival of haplogroup N and gradual replacement of R1a lineages in North-Eastern Europe