Open access Investigating Holocene human population history in North Asia using ancient mitogenomes, by Kılınç et al., Scientific Reports (2018) 8: 8969.
Abstract (emphasis mine):
Archaeogenomic studies have largely elucidated human population history in West Eurasia during the Stone Age. However, despite being a broad geographical region of significant cultural and linguistic diversity, little is known about the population history in North Asia. We present complete mitochondrial genome sequences together with stable isotope data for 41 serially sampled ancient individuals from North Asia, dated between c.13,790 BP and c.1,380 BP extending from the Palaeolithic to the Iron Age. Analyses of mitochondrial DNA sequences and haplogroup data of these individuals revealed the highest genetic affinity to present-day North Asian populations of the same geographical region suggesting a possible long-term maternal genetic continuity in the region. We observed a decrease in genetic diversity over time and a reduction of maternal effective population size (Ne) approximately seven thousand years before present. Coalescent simulations were consistent with genetic continuity between present day individuals and individuals dating to 7,000 BP, 4,800 BP or 3,000 BP. Meanwhile, genetic differences observed between 7,000 BP and 3,000 BP as well as between 4,800 BP and 3,000 BP were inconsistent with genetic drift alone, suggesting gene flow into the region from distant gene pools or structure within the population. These results indicate that despite some level of continuity between ancient groups and present-day populations, the region exhibits a complex demographic history during the Holocene.
Interesting excerpts:
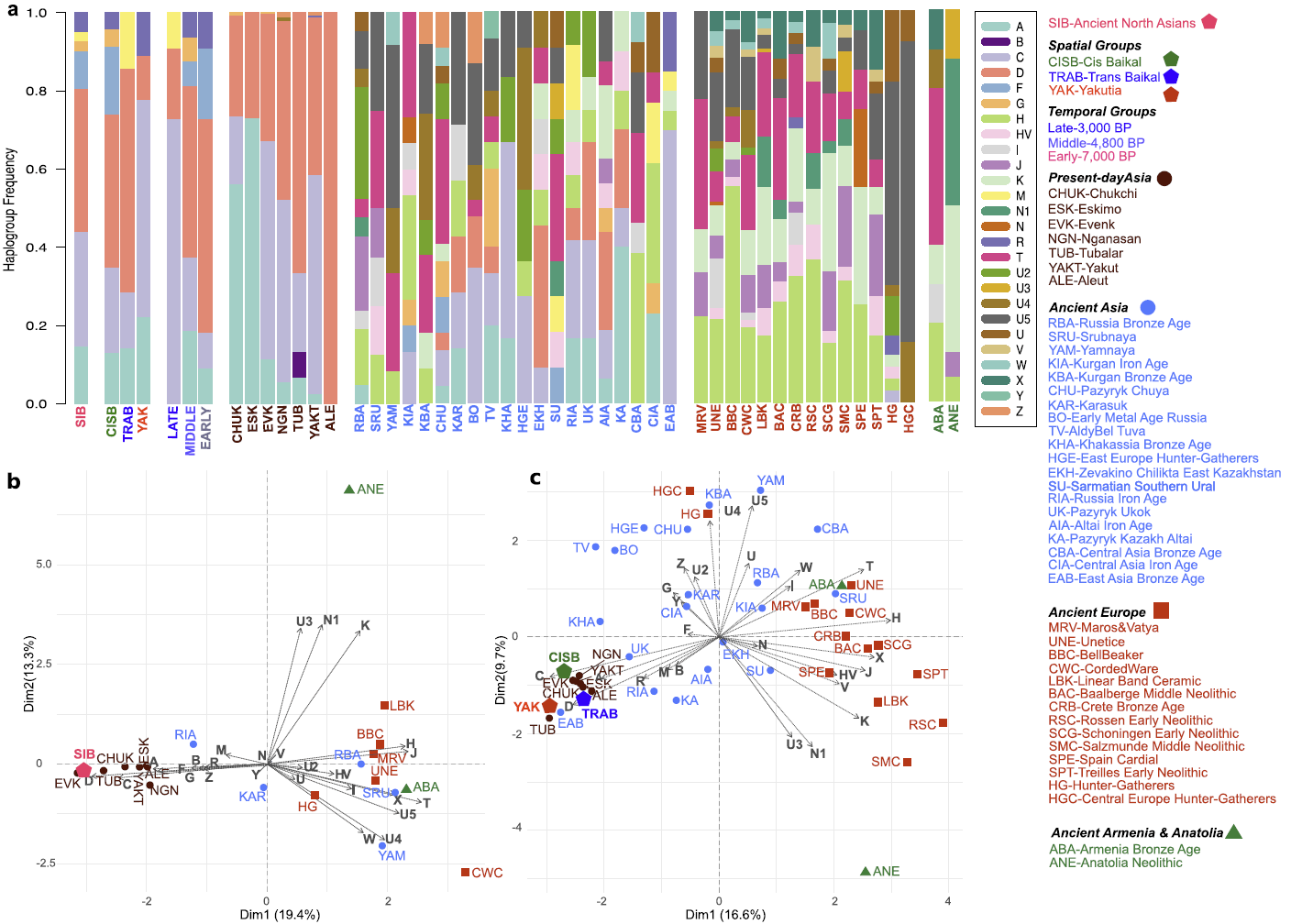
Although highly dependent on sample size and thus prone to generalization, haplotype sharing analysis between three spatial groups and other modern and ancient populations (Supplementary Table S15) revealed that the TRAB group shared most lineages with ancient Kazakh Altai (KA) and modern Nganasan (NGN)39,40,41,42. The CISB group shared most lineages with Tubalar39,42, KA43 and Early Bronze Age groups of Russia (BO)12, which might reflect the Siberian roots of BO, consistent with MDS based on Fst (Fig. 3b). The YAK group shared most lineages with the CISB, BO and Tubalar groups. These results showed that despite being from different sides of the Lake Baikal, the CISB and YAK groups shared most lineages with the Tubalar and also both of them were to a certain degree affiliated to the BO of the Cis-Baikal region, thus, reflecting a shared common ancestry. Furthermore, the CISB and YAK groups share lineages supporting the hypothesis of a lasting continuity in this large geographical territory. However, the TRAB group may have different legacy with affinities to ancient Kazakh Altai and modern Nganasan groups (that, actually, may have relocated from the Trans-Baikal region in times post-dating our sample).
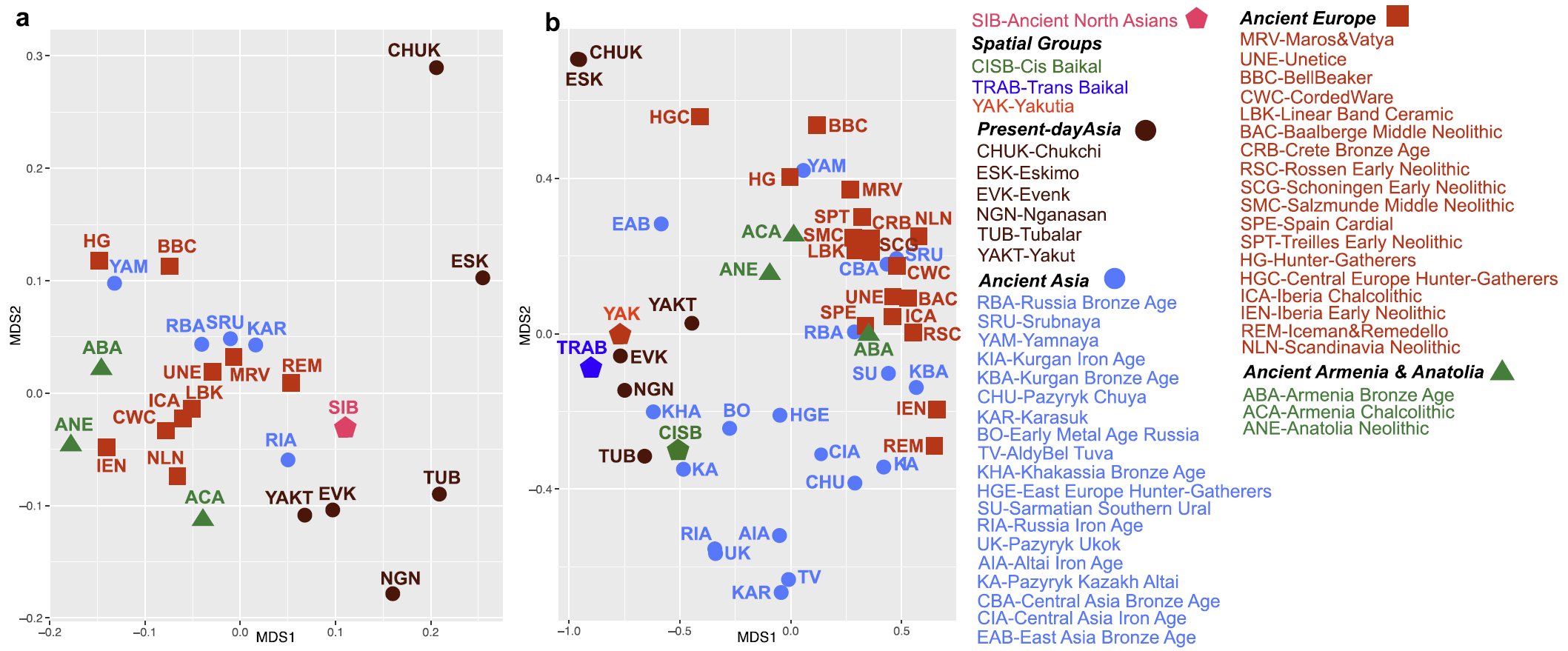
Two findings, however, were intriguing. One was the discovery of only weak support for a single regional population in comparisons between Early vs. Late as well as Middle vs. Late groups in the region. This may be explained by population structure, as the Late group comprised geographically very distant individuals, such as individuals from Krasnoyarsk Krai and Amur Oblast, not represented in the other diachronic groups (Table S9). Another explanation for rejecting the null hypothesis of continuity between the Middle and Late (4,800–3,000 BP) groups might be due to an interruption and the arrival of pastoralists at the beginning of the Iron Age between 3,670 to 2,760 BP as suggested by the archaeological record32. Thus, the introduction of the new lifeways, technologies and material culture expressions might also here be associated to an increased mobility into the area.
The second point was the estimated reduction in maternal effective population size and haplotype diversity around 7,000 BP. Intriguingly, climate modelling and radiocarbon dating studies53 suggest that climatic change and a collapse of the riverine ecosystems might have affected the human populations in Cis Baikal between 7,000–6,000 BP in line with our results. This finding was further supported by archaeological studies pointing to a possible hiatus38,54,55.
Although our results provide a first glimpse into population structure and diversity in North Asia during the Holocene which link to trend in the archaeological record, complete genome sequences will provide a higher resolution of more complex demographic events in the region.
Yet another hint at the west-east (and not east-west) population movement in Eurasia after the Corded Ware and Yamna expansions, without any significant change in the other direction until the Iron Age (as we know from Fennoscandian samples), which leaves still less space to propose incoming Uralic-speaking groups from Asia…
Related:
- Hungarian mitogenomes similar to East and West Slavs, but genetic substratum predates their historic contacts
- Pre-Germanic born out of a Proto-Finnic substrate in Scandinavia
- Consequences of Damgaard et al. 2018 (III): Proto-Finno-Ugric & Proto-Indo-Iranian in the North Caspian region
- On Proto-Finnic language guesstimates, and its western homeland
- Oldest N1c1a1a-L392 samples and Siberian ancestry in Bronze Age Fennoscandia
- The concept of “Outlier” in Human Ancestry (III): Late Neolithic samples from the Baltic region and origins of the Corded Ware culture
- Genetic prehistory of the Baltic Sea region and Y-DNA: Corded Ware and R1a-Z645, Bronze Age and N1c
- Uralic as a Corded Ware substrate of Indo-Iranian, and loanwords in Finno-Ugric
- More evidence on the recent arrival of haplogroup N and gradual replacement of R1a lineages in North-Eastern Europe