New preprint The population history of northeastern Siberia since the Pleistocene, by Sikora et al. bioRxiv (2018).
Interesting excerpts (emphasis mine; most internal references removed):
ANE ancestry
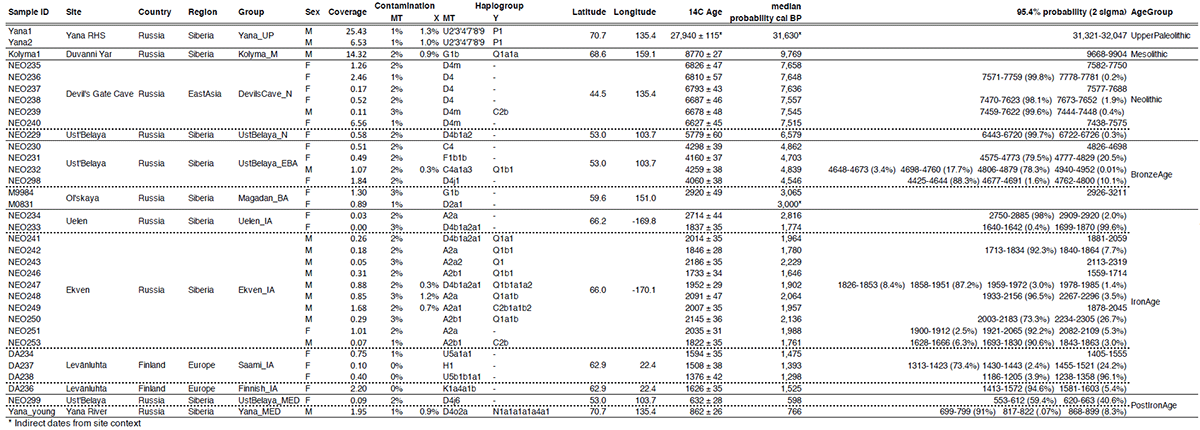
The earliest, most secure archaeological evidence of human occupation of the region comes from the artefact-rich, high-latitude (~70° N) Yana RHS site dated to ~31.6 kya (…)
The Yana RHS human remains represent the earliest direct evidence of human presence in northeastern Siberia, a population we refer to as “Ancient North Siberians” (ANS). Both Yana RHS individuals were unrelated males, and belong to mitochondrial haplogroup U, predominant among ancient West Eurasian hunter-gatherers, and to Y chromosome haplogroup P1, ancestral to haplogroups Q and R, which are widespread among present-day Eurasians and Native Americans.
Symmetry tests using f4 statistics reject tree-like clade relationships with both Early West Eurasians (EWE; Sunghir) and Early East Asians (EEA; Tianyuan); however, Yana is genetically closer to EWE, despite its geographic location in northeastern Siberia
Using admixture graphs (qpGraph) and outgroup-based estimation of mixture proportions (qpAdm), we find that Yana can be modelled as EWE with ~25% contribution from EEA
Among all ancient individuals, Yana shares the most genetic drift with Mal’ta, and f4 statistics show that Mal’ta shares more alleles with Yana than with EWE (e.g. f4(Mbuti,Mal’ta;Sunghir,Yana) = 0.0019, Z = 3.99). Mal’ta and Yana also exhibit a similar pattern of genetic affinities to both EWE and EEA, consistent with previous studies.The ANE lineage can thus be considered a descendant of the ANS lineage, demonstrating that by 31.6 kya early representatives of this lineage were widespread across northern Eurasia, including far northeastern Siberia.
Ancient Palaeosiberian
(…) the 9.8 kya Kolyma1 individual, representing a group we term “Ancient Paleosiberians” (AP). Our results indicate that AP are derived from a first major genetic shift observed in the region. Principal component analysis (PCA), outgroup f3-statistics and mtDNA and Y chromosome haplogroups (G1b and Q1a1a, respectively) demonstrate a close affinity between AP and present-day Koryaks, Itelmen and Chukchis, as well as with Native Americans.
For both AP and Native Americans, ANS ancestry appears more closely related to Mal’ta than Yana, therefore rejecting a direct contribution of Yana to later AP or Native American groups.
Lake Baikal Neolithic – Bronze Age
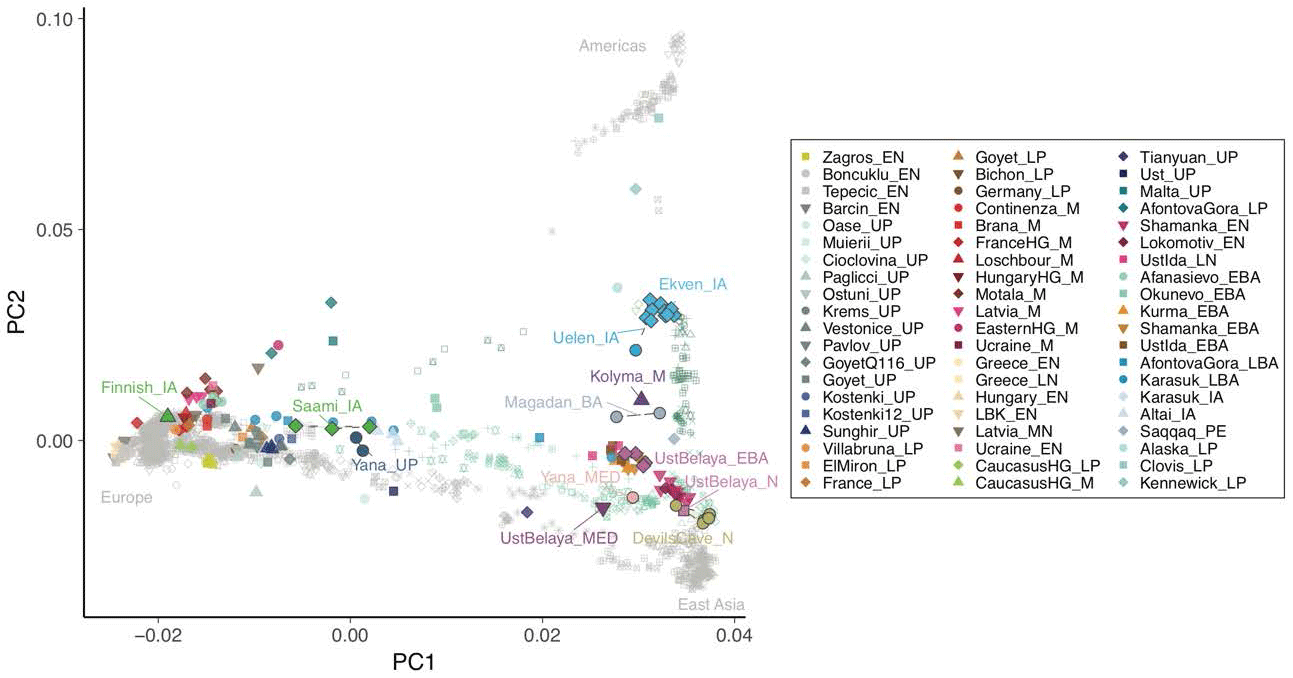
(…) the newly reported genomes from Ust’Belaya and recently published neighbouring Neolithic and Bronze Age sites show a succession of three distinct genetic ancestries over a ~6 ky time span. The earliest individuals show predominantly East Asian ancestry, closely related to the ancient individuals from DGC. In the early Bronze Age (BA), we observe a resurgence of AP ancestry (up to ~50% ancestry fraction), as well as influence of West Eurasian Steppe ANE ancestry represented by the early BA individuals from Afanasievo in the Altai region (~10%) This is consistent with previous reports of gene flow from an unknown ANE-related source into Lake Baikal hunter-gatherers.
Our results suggest a southward expansion of AP as a possible source, which is also consistent with the replacement of Y chromosome lineages observed at Lake Baikal, from predominantly haplogroup N in the Neolithic to haplogroup Q in the BA. Finally, the most recent individual from Ust’Belaya, dated to ~600 years ago, falls along the Neosiberian cline, similar to the ~760 year-old ‘Young Yana’ individual from northeastern Siberia, demonstrating the widespread distribution of Neosiberian ancestry in the most recent epoch.
Finland Saami
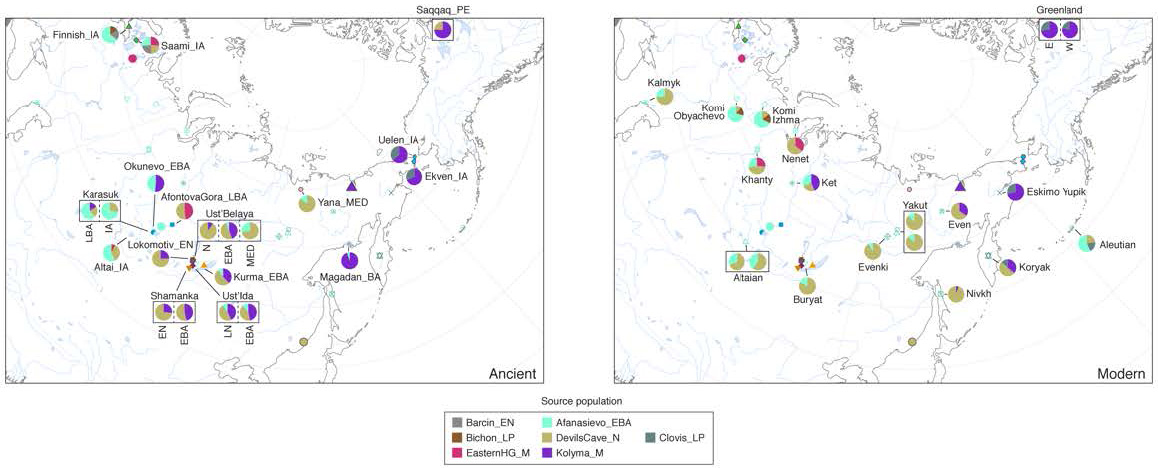
At the western edge of northern Eurasia, genetic and strontium isotope data from ancient individuals at the Levänluhta site documents the presence of Saami ancestry in Southern Finland in the Late Holocene 1.5 kya. This ancestry component is currently limited to the northern fringes of the region, mirroring the pattern observed for AP ancestry in northeastern Siberia. However, while the ancient Saami individuals harbour East Asian ancestry, we find that this is better modelled by DGC rather than AP, suggesting that AP influence was likely restricted to the eastern side of the Urals. Comparison of ancient Finns and Saami with their present-day counterparts reveals additional gene flow over the past 1.6 kya, with evidence for West Eurasian admixture into modern Saami. The ancient Finn from Levänluhta shows lower Siberian ancestry than modern Finns .
EDIT (27 OCT 2018): By comparing the three, I see these are samples published already (at least two) in Lamnidis et al. (2018), but here with added (1) specific radiocarbon dates, (2) comparison with Neosiberian populations and (3) strontium isotope analyses.
Finnish_IA (ca. 350 AD) is probably a Saami-speaking individual, just like the Saami_IA with newly reported radiocarbon dates from Levänluhta ca. 400-600 AD (since Fennic peoples were then likely around the Gulf of Finland).
The conflicting strontium isotope data on marine dietary resources on certain samples from the supplementary material hint at possible external origin of the diet of some of the previously reported (and possibly one newly reported) Saami Iron Age individuals, from some 25-30 km. to the northwest through the river up to hundreds of km. to the southwest of Levänluhta (i.e. the whole coast of the Bothnian Sea). It is unclear why they would prefer an origin of the dietary source in southern Baltic regions instead of some km. to the west, though, unless that’s what they want to propose based on the sample’s admixture…
The coast of the Bothnian Sea (=the northern part of the Baltic Sea, between Sweden and Finland) lay only 25-30 km to the northwest, and accessible to the Iron Age people of the Levänluhta region via the Kyrönjoki river. (…) For individual JA2065/DA236, the low 87Sr/86Sr value (0.71078) would imply an exceptionally heavy reliance on Baltic Sea resources. The δ13C and δ15N values of the individual are near comparable (especially considering within-Baltic latitudinal gradients in δ13C; Torniainen et al. 2017) to the δ13C and δ15N values of a Middle Neolithic population on the Baltic island of Gotland (Eriksson, 2004) interpreted to have subsisted primarily on seals.
These new data on the samples give us some more information than what we already had, because the early date of Finnish_IA implies that there was few East Asian admixture (if any at all) in west Finland during the Roman Iron Age, which pushes still farther forward in time the expected appearance of Siberian ancestry among Saamic (first) and Fennic populations (later). It is unclear whether this East Asian ancestry found in Finnish_IA is actually related to DGC, or it is rather related to the ENA-like ancestry found already in Baltic hunter-gatherers (i.e. in some EHG samples from Karelia), for which Baikal_EN is a good proxy in Lazaridis et al. (2018).
Since Bronze Age and Iron Age samples from Estonia show more Baltic_HG drift compared to Corded Ware samples, it is likely that this supposedly DGC-related ancestry (here considered part of the ‘Siberian ancestry’) is actually an EHG-related ENA component of north-east European hunter-gatherers, with whom Finno-Saamic peoples admixed during the expansion of the Corded Ware culture into Finland.
The paper finds thus increased (probably the actual) Siberian ancestry in modern Finns compared to this Iron Age Saami individual. Coupled with the later Saami Iron Age samples, from between one to three centuries later – showing the start of Siberian ancestry influx – , we can begin to establish when the expansion of Siberian ancestry happened in central Finland, and thus quite likely when the Saami began to expand to the north and east and admix with Palaeo-Laplandic peoples.
One sample of haplogroup N1a1a1a1a4a1-M1982, Yana_MED, is found in the Arctic region (north-eastern Yakutia) ca. 1100 AD. Since it is derived from N1a1a1a1a-L392, it might be a surprise for some to find it in a clearly non-Uralic speaking environment at the same time other subclades of this haplogroup were admixing in the west with well-established Finno-Saamic, Volga-Finnic, Ugric, and Samoyedic populations…
On the growing doubts that these data – contradicting the CWC=IE theory – are creating among geneticists (from the supplementary materials):
NOTE. This paper comes from the Copenhagen group, also signed by Kristiansen, one of today’s strongest supporters of this connection…
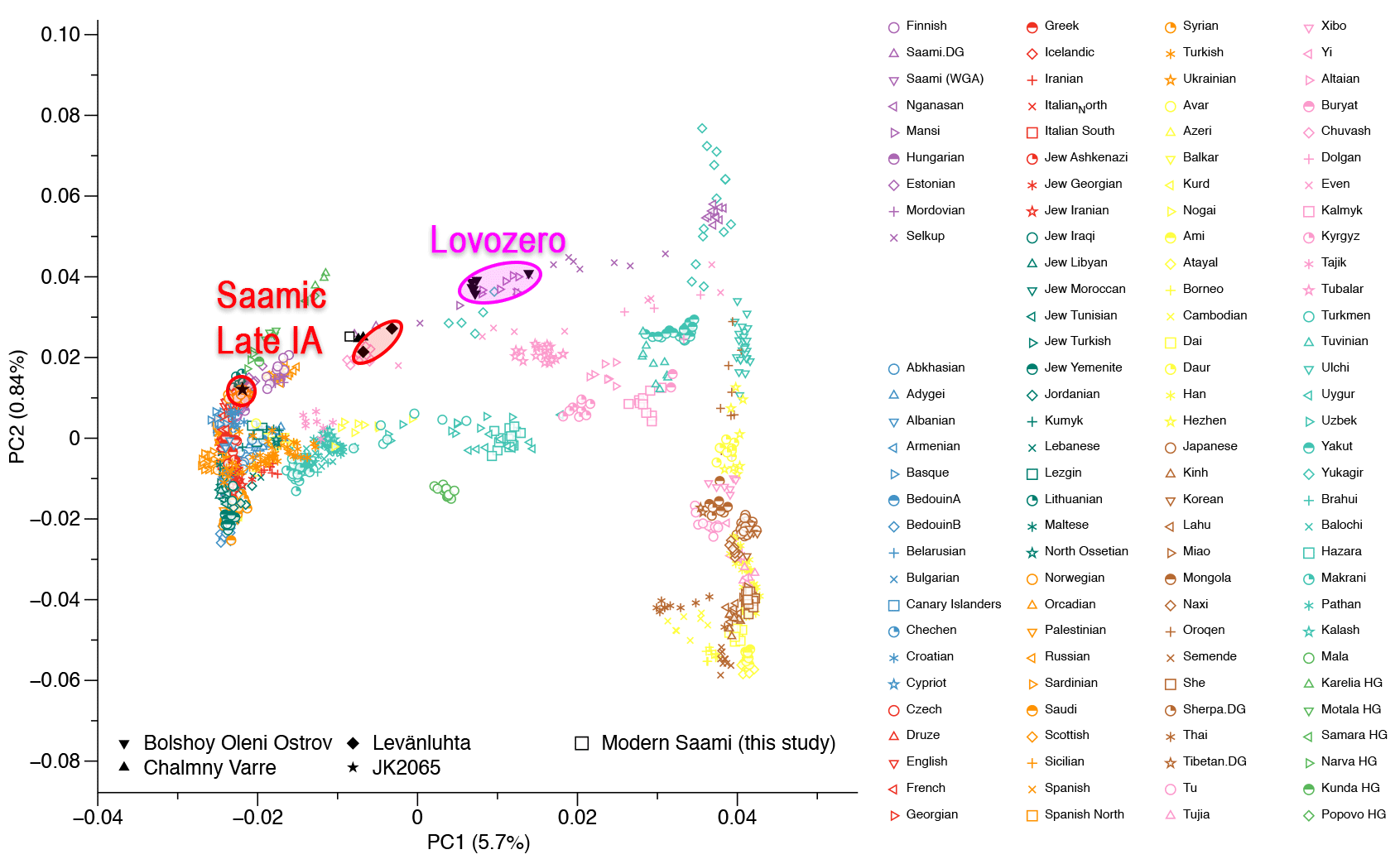
The Proto-Saami language evolved in southern Finland and Karelia in the Early Iron Age, an area now host to Finnish and the closely related Karelian, but with Saami toponyms showing that the latter two languages are intrusive here (Saarikivi 2004). Saami-speaking populations are thought to have retreated to Lapland during the Middle Iron Age (300–800 AD), where it diverged into the modern Saami dialects. Genetically, the northward retreat of the Saami language correlates with the documented decrease of Saami ancestry in Southern Finland between the Iron Age and the modern period (cf. Lamnidis et al. 2018).
On the way to Lapland, the Saami replaced at least two linguistically obscure groups. This can be inferred from 1) an influx of non-Uralic loanwords into Proto-Saami in the Finnish Lakeland area, and 2) an influx of non-Uralic, non-Germanic words into Saami dialects in Lapland (Aikio 2012). Both of these borrowing events imply contact with non-Saami-speaking groups, e.g. non-Uralic-speaking hunter-gatherers that may have left a genetic and linguistic footprint on modern Saami populations.
The linguistic prehistory of Finland thus does not allow for a straightforward interpretation of the genetic data. The detection of East Asian ancestry in the genetically Saami individual is indicative of a population movement from the east (cf. Lamnidis et al. 2018, Rootsi et al. 2007), one that given the affinities with the ~7.6 ky old individuals from the Devil’s Gate Cave may have been a western extension of the Neosiberian turnover. However, it remains unclear whether this gene flow should be associated with the arrival of Uralic speakers, thus providing further support for a Uralic homeland in Eastern Eurasia, or with an earlier immigration of pre-Uralic, so-called “Paleo-Lakelandic” groups.
I think the genetic interpretation is already straightforward, though. We had a sneak peek at how this late admixture with non-Uralians (mainly Palaeo-Lakelandic and Palaeo-Laplandic peoples from Lovozero and related asbestos ware cultures) is going to unfold among expanding Saami-speaking populations thanks to Lamnidis et al. (2018):
Also, still no trace of R1a in far East Asia (reported as M17 ca. 5300 BC near Lake Baikal by Moussa et al. 2016), so I still have doubts about my previous assessment that R1a split into M17 (and thus also M417) in Siberia, with those expanding hunter-gatherer pottery.
Related
- Dzudzuana, Sidelkino, and the Caucasus contribution to the Pontic-Caspian steppe
- Updated phylogenetic tree of haplogroup Q-M242 points to Palaeolithic expansions
- Iron Age bottleneck of the Proto-Fennic population in Estonia
- Corded Ware—Uralic (II): Finno-Permic and the expansion of N-L392/Siberian ancestry
- Corded Ware—Uralic (I): Differences and similarities with Yamna
- Haplogroup R1a and CWC ancestry predominate in Fennic, Ugric, and Samoyedic groups
- On the origin and spread of haplogroup R1a-Z645 from eastern Europe
- Oldest N1c1a1a-L392 samples and Siberian ancestry in Bronze Age Fennoscandia
- Consequences of Damgaard et al. 2018 (III): Proto-Finno-Ugric & Proto-Indo-Iranian in the North Caspian region
- Steppe and Caucasus Eneolithic: the new keystones of the EHG-CHG-ANE ancestry in steppe groups