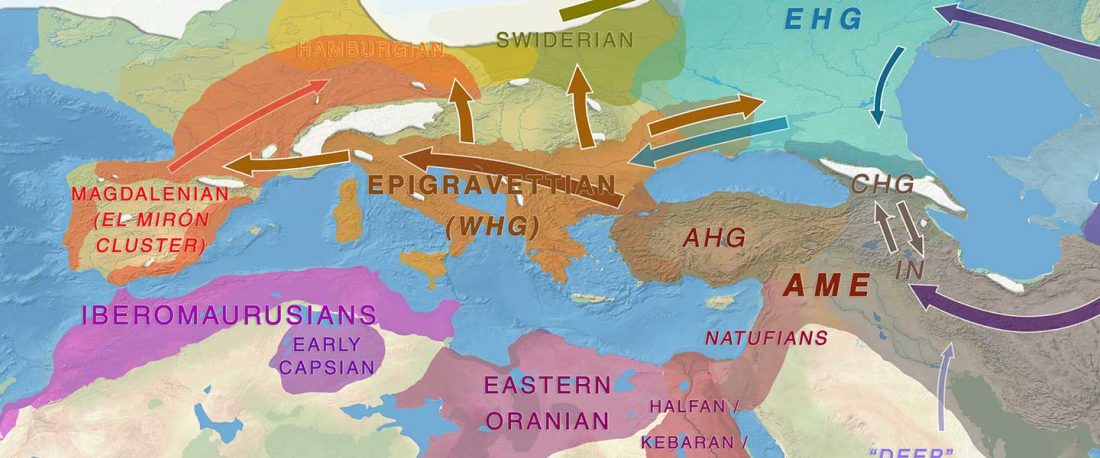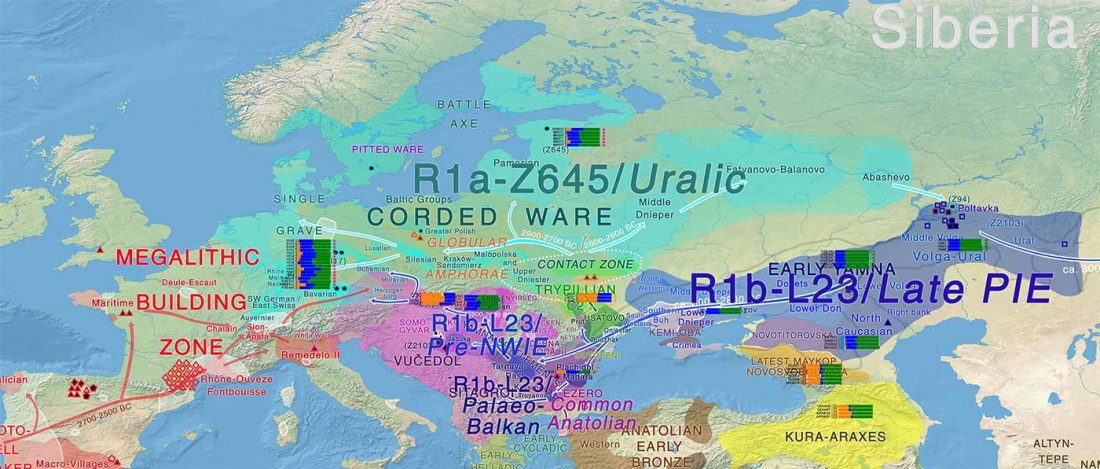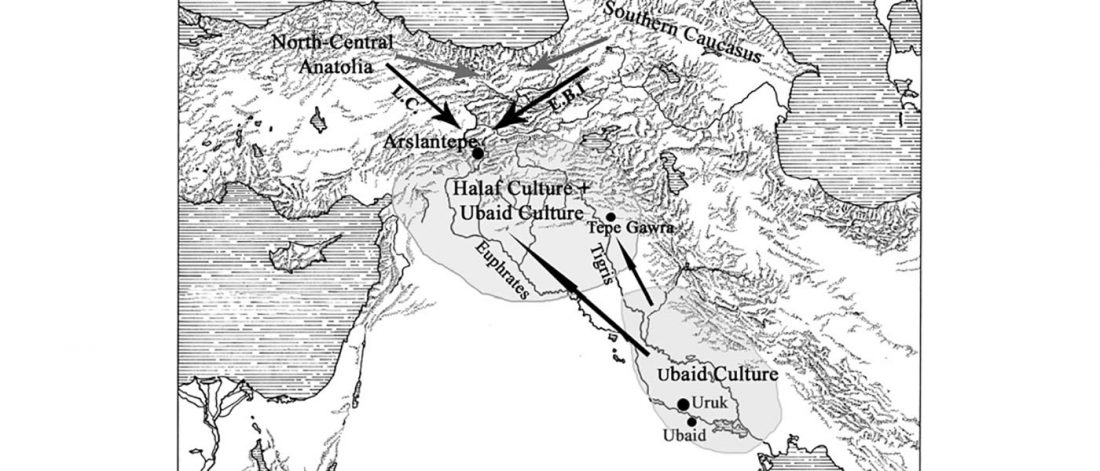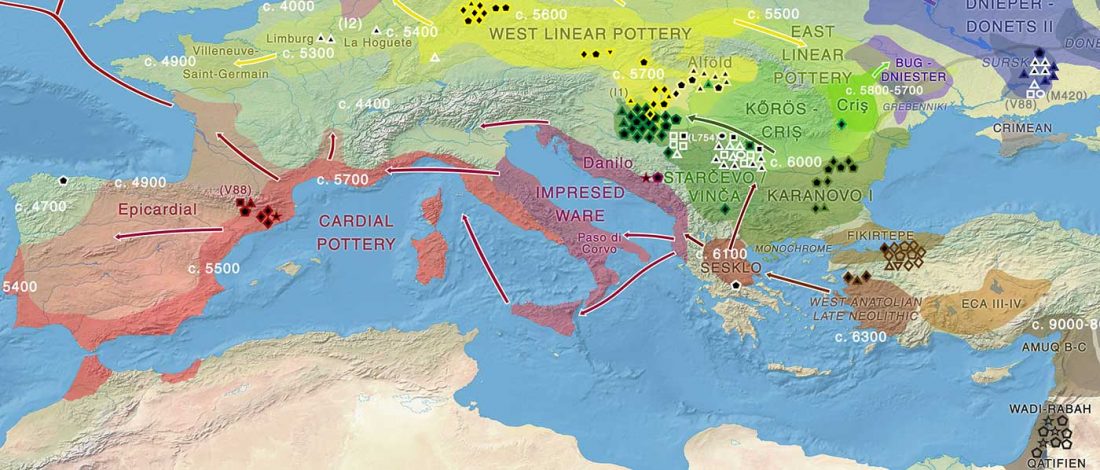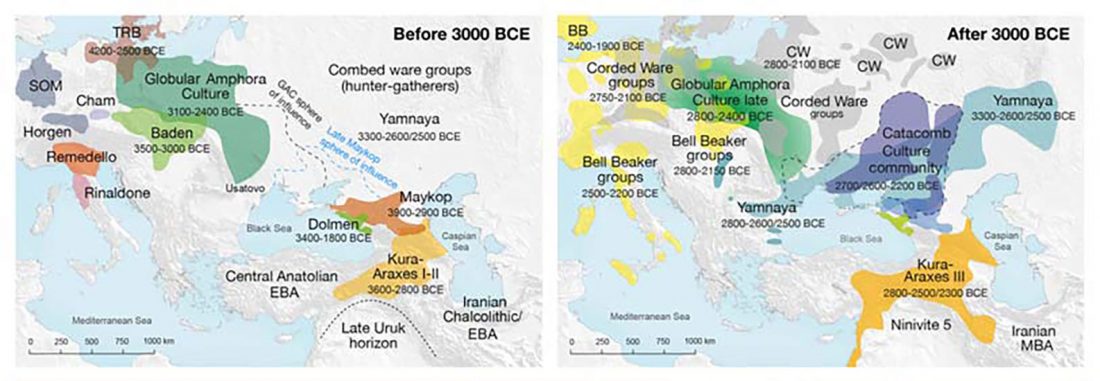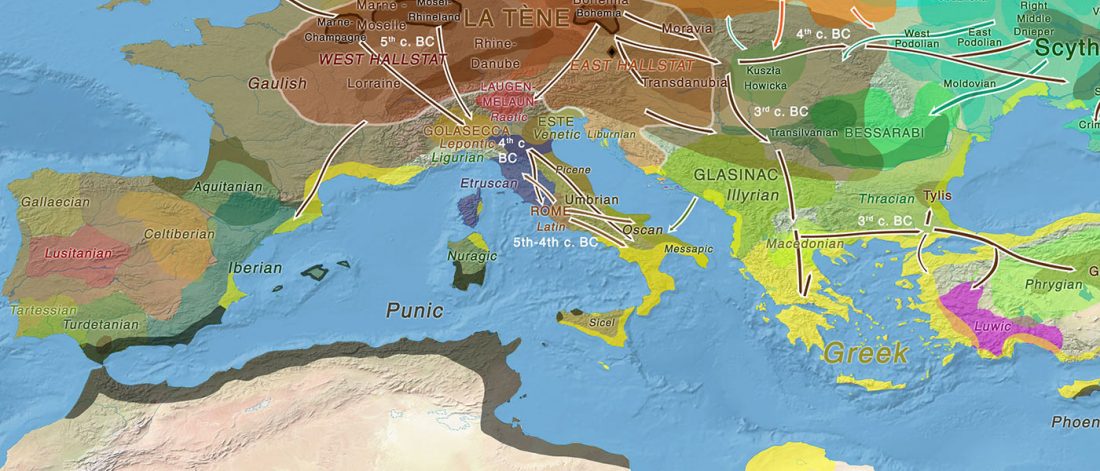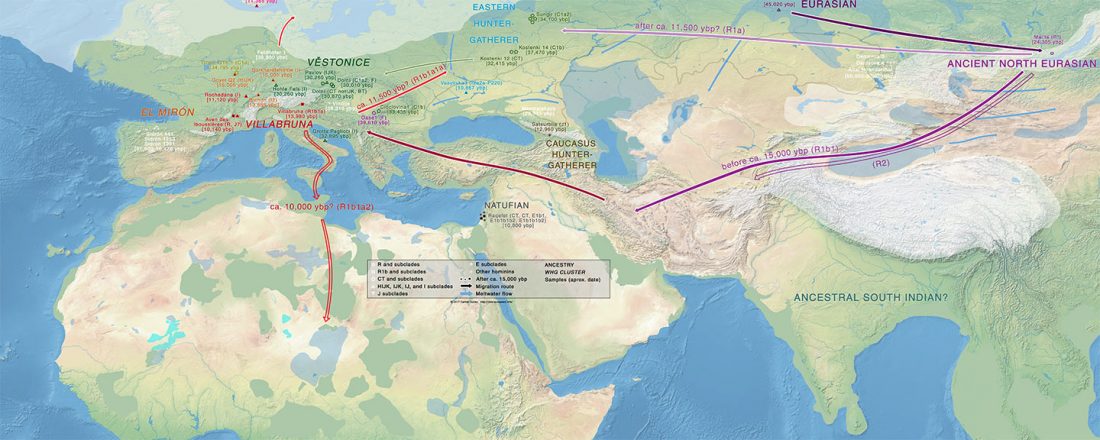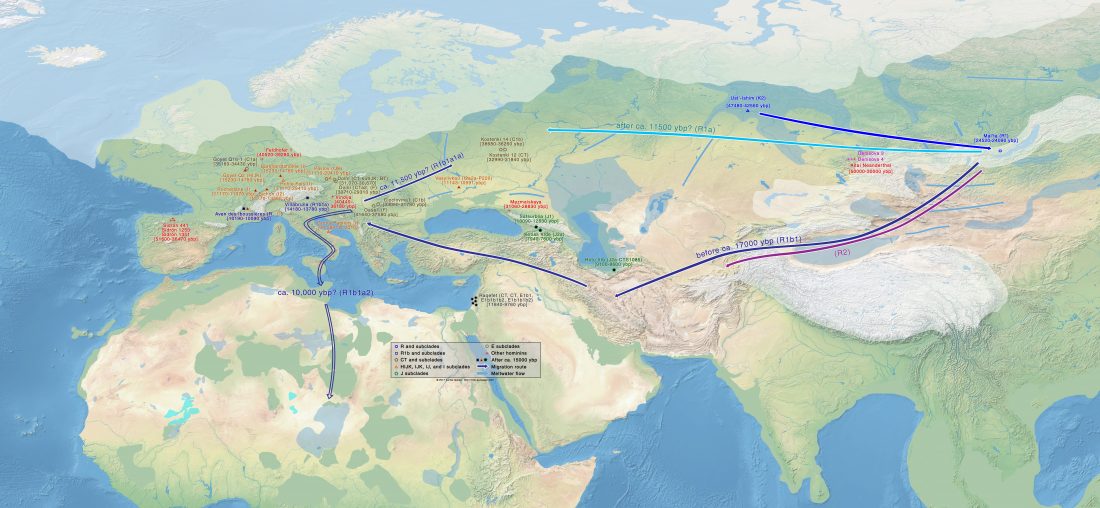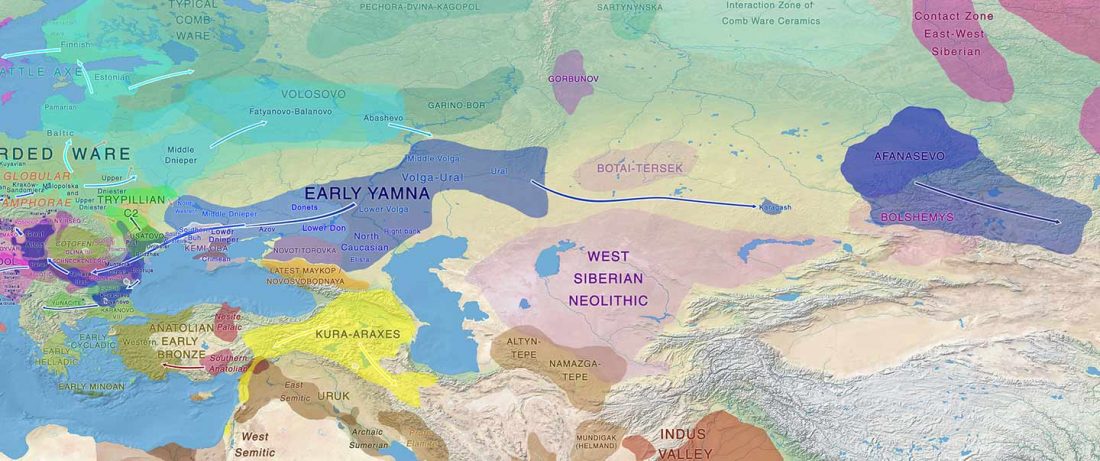
New paper (behind paywall) by David Anthony, Archaeology, Genetics, and Language in the Steppes: A Comment on Bomhard, complementing in a favourable way Bomhard’s Caucasian substrate hypothesis in the current issue of the JIES.
NOTE. I have tried to access this issue for some days, but it’s just not indexed in my university library online service (ProQuest) yet. This particular paper is on Academia.edu, though, as are Bomhard’s papers on this issue in his site.
Interesting excerpts (emphasis mine):
… Read the rest “Volga Basin R1b-rich Proto-Indo-Europeans of (Pre-)Yamnaya ancestry”Along the banks of the lower Volga many excavated hunting-fishing camp sites are dated 6200-4500 BC. They