Open access Prehistoric migrations through the Mediterranean basin shaped Corsican Y-chromosome diversity, by Di Cristofaro et al. PLOS One (2018).
Interesting excerpts:
This study included 321 samples from men throughout Corsica; samples from Provence and Tuscany were added to the cohort. All samples were typed for 92 Y-SNPs, and Y-STRs were also analyzed.
Haplogroup R represented approximately half of the lineages in both Corsican and Tuscan samples (respectively 51.8% and 45.3%) whereas it reached 90% in Provence. Sub-clade R1b1a1a2a1a2b-U152 predominated in North Corsica whereas R1b1a1a2a1a1-U106 was present in South Corsica. Both SNPs display clinal distributions of frequency variation in Europe, the U152 branch being most frequent in Switzerland, Italy, France and Western Poland. Calibrated branch lengths from whole Y chromosome sequencing [44,45] and ancient DNA studies [46] both indicated that R1a and R1b diversification began relatively recently, about 5 Kya, consistent with Bronze Age and Copper Age demographic expansion. TMRCA estimations are concordant with such expansion in Corsica.
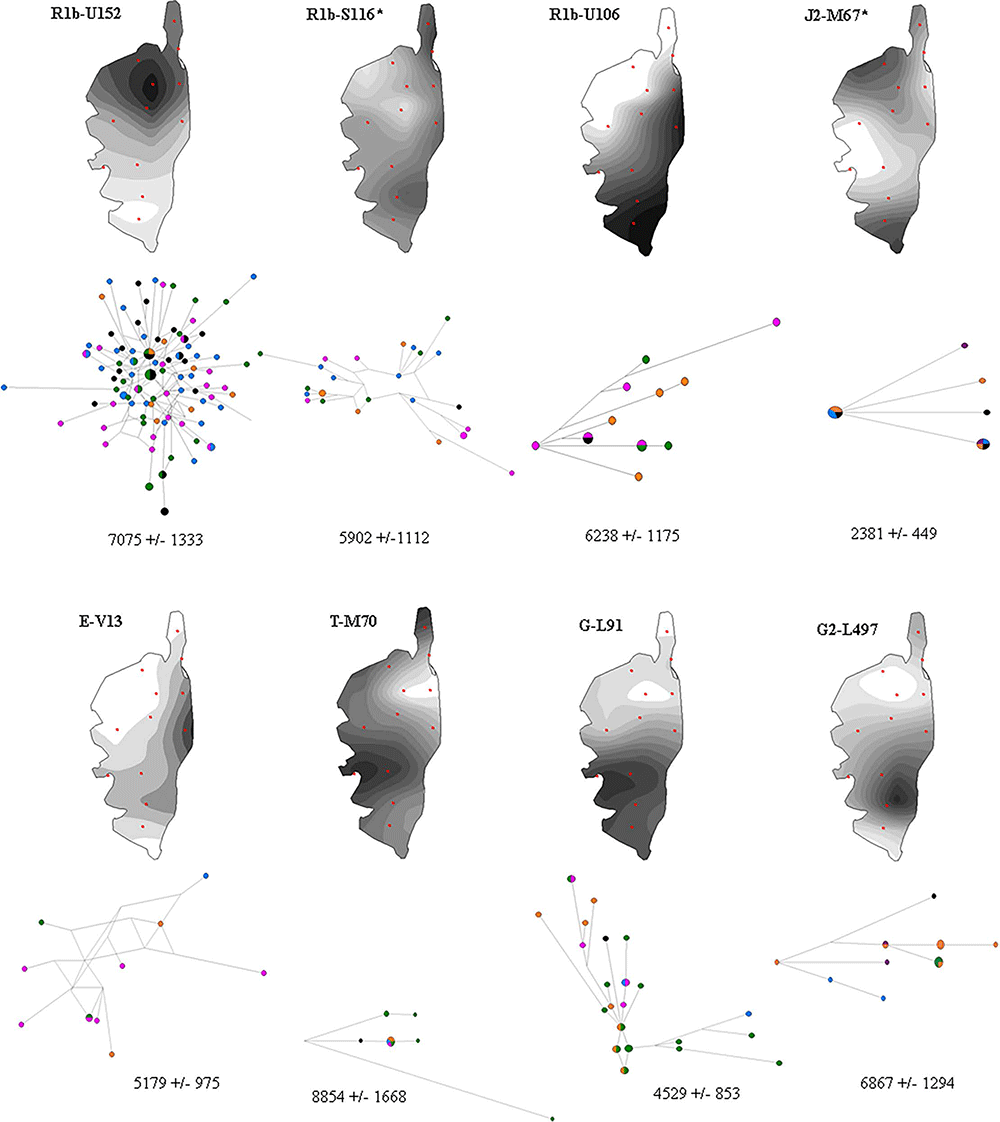
Haplogroup G reached 21.7% in Corsica and 13.3% in Tuscany. Sub-clade G2a2a1a2-L91 accounted for 11.3% of all haplogroups in Corsica yet was not present in Provence or in Tuscany. Thirty-four out of the 37 G2a2a1a2-L91 displayed a unique Y-STR profile, illustrated by the star-like profile of STR networks (Fig 1). G2a2a1a2-L91 and G2a2a-PF3147(xL91xM286) show their highest frequency in present day Sardinia and southern Corsica compared to low levels from Caucasus to Southern Europe, encompassing the Near and Middle East [21,47–50]. Ancient DNA results from Early and Middle Neolithic samples reported the presence of haplogroup G2a-P15 [51–53], consistent with gene flow from the Mediterranean region during the Neolithic transition. Td expansion time estimated by STR for P15-affiliated chromosomes was estimated to be 15,082+/-2217 years ago [49]. Ötzi, the 5,300-year-old Alpine mummy, was derived for the L91 SNP [21]. A genetic relationship between G haplogroups from Corsica and Sardinia is further supported by DYS19 duplication, reported in North Sardinia [14], and observed in the southern part of the Corsica in 9 out of 37 G2a2a1a2-L91 chromosomes and in 4 out of 5 G2a2a-PF3147(xL91xM286) chromosomes, 3 of which displayed an identical STR profile (S4 Table).
This lineage has a reported coalescent age estimated by whole sequencing in Sardinian samples of about 9,000 years ago. This could reflect common ancestors coming from the Caucasus and moving westward during the Neolithic period [48], whereas their continental counterparts would have been replaced by rapidly expanding populations associated with the Bronze Age [46,54,55]. Estimated TMRCA for L91 lineage in Corsica is 4529 +/- 853 years. G-L497 showed high frequencies in Corsica compared to Provence and Tuscany, and this haplogroup was common in Europe, but rare in Greece, Anatolia and the Middle East. Fifteen out of the 17 Corsican G2a2b2a1a1b-L497 displayed a unique Y-STR profile (S4 Table) with an estimated TMRCA of 6867 +/- 1294 years. Haplogroup G2a2b1-M406, associated with Impressed Ware Neolithic markers, along with J2a1-DYS445 = 6 and J2a1b1-M92 [22,49], had very low levels in Corsica. Conversely, G2a2b2a-P303was highly represented and seemed to be independent of the G2a2b1-M406 marker. The 7 G2a2b2a-P303(xL497xM527) Corsican chromosomes displayed a unique Y-STR profile (S4 Table).
Haplogroup J, mainly represented by J2a1b-M67(xM92), displayed intermediate frequencies in Corsica compared to Tuscany and Provence. J2a1b-M67(xM92) derived STR network analysis displayed a quite homogeneous profile across the island with an estimated TMRCA of 2381 +/- 449 years (Fig 1) and individuals displaying M67 were peripheral compared to Northwestern Italians (S2 Fig). The haplogroup J2a1-Page55(xM67xM530), characteristic of non-Greek Anatolia [22], was found in the north-west of Corsica. Haplogroup J2a1-DYS445 = 6 was found in the north-west with DYS391 = 10 repeats, and in the far south with DYS391 = 9 repeats, the former was associated with Anatolian Greek samples, whereas the second was found in central Anatolia [22]. The 7 J2b2a-M241 displayed a unique Y-STR profile (S4 Table), they were only detected in the Cap Corse region, this sub-haplogroup shows frequency peaks in both the southern Balkans and northern-central Italy [56] and is associated with expansion from the Near East to the Balkans during Neolithic period [57].
Haplogroup E, mainly represented by E1b1b1a1b1a-V13, displayed intermediate frequencies in Corsica compared to Tuscany and Provence. E1b1b1a1b1a-V13 was thought to have initiated a pan-Mediterranean expansion 7,000 years ago starting from the Balkans [52] and its dispersal to the northern shore of the Mediterranean basin is consistent with the Greek Anatolian expansion to the western Mediterranean [22], characteristic of the region surrounding Alaria, and consistent with the TMRCA estimated in Corsica for this haplogroup. A few E1b1a-V38 chromosomes are also observed in the same regions as V13.
Related:
- Olalde et al. and Mathieson et al. (Nature 2018): R1b-L23 dominates Bell Beaker and Yamna, R1a-M417 resurges in East-Central Europe during the Bronze Age
- Haplogroup J spread in the Mediterranean due to Phoenician and Greek colonizations
- Migration vs. Acculturation models for Aegean Neolithic in Genetics — still depending strongly on Archaeology
- Ancient Phoenician mtDNA from Sardinia, Lebanon reflects settlement, genetic diversity, and female mobility
- Genetic origins of Minoans and Mycenaeans and their continuity into modern Greeks