New preprint The Arrival of Steppe and Iranian Related Ancestry in the Islands of the Western Mediterranean by Fernandes, Mittnik, Olalde et al. bioRxiv (2019)
Interesting excerpts (emphasis in bold; modified for clarity):
Balearic Islands: The expansion of Iberian speakers
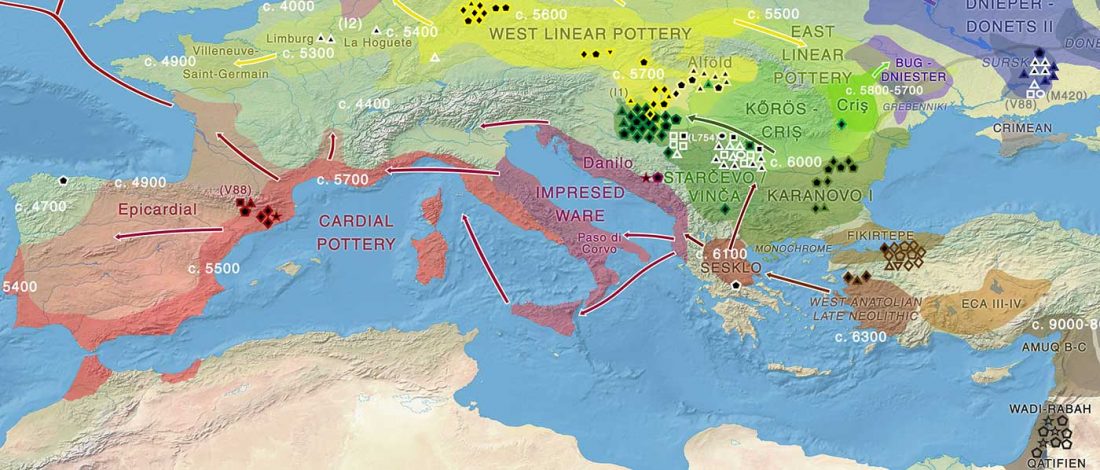
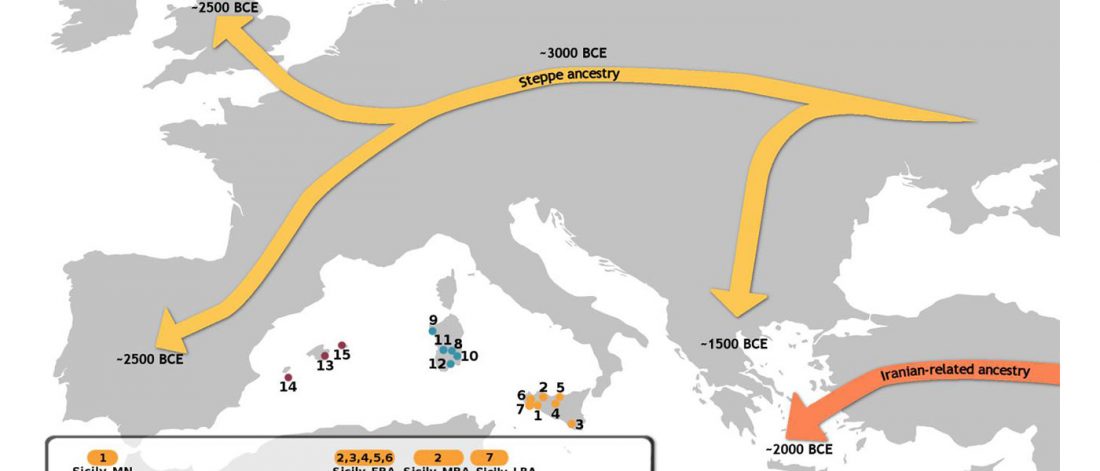
… Read the rest “Arrival of steppe ancestry with R1b-P312 in the Mediterranean: Balearic Islands, Sicily, and Iron Age Sardinia”Mallorca_EBA dates to the earliest period of permanent occupation of the islands at around 2400 BCE. We parsimoniously modeled Mallorca_EBA as deriving 36.9 ± 4.2% of her ancestry from a source related to Yamnaya_Samara; (…). We next used qpAdm to identify “proximal” sources for Mallorca_EBA’s ancestry that are more closely related to this individual in space and time, and found