Content under CC-BY-NC license. Interesting excerpts (emphasis mine):
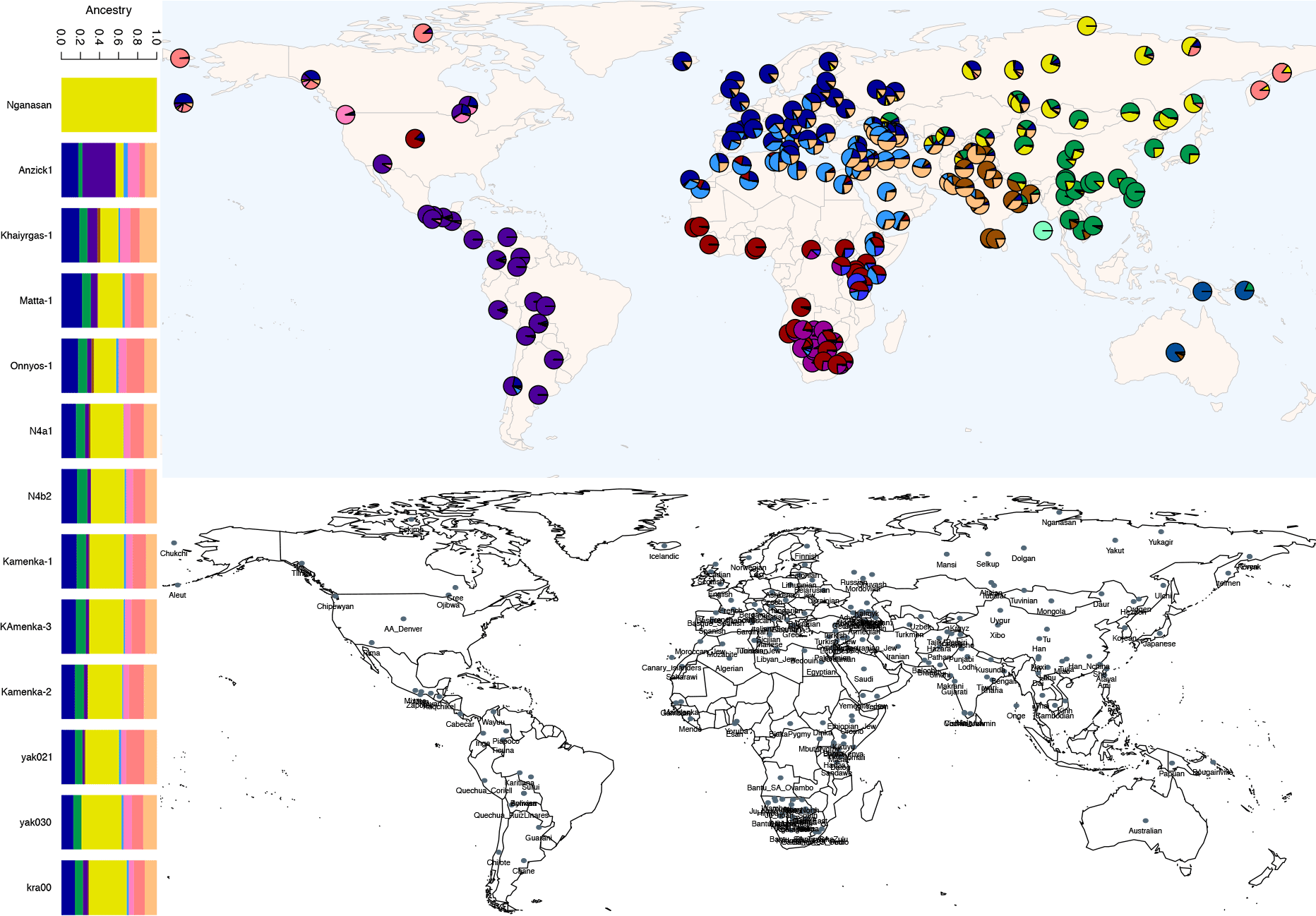
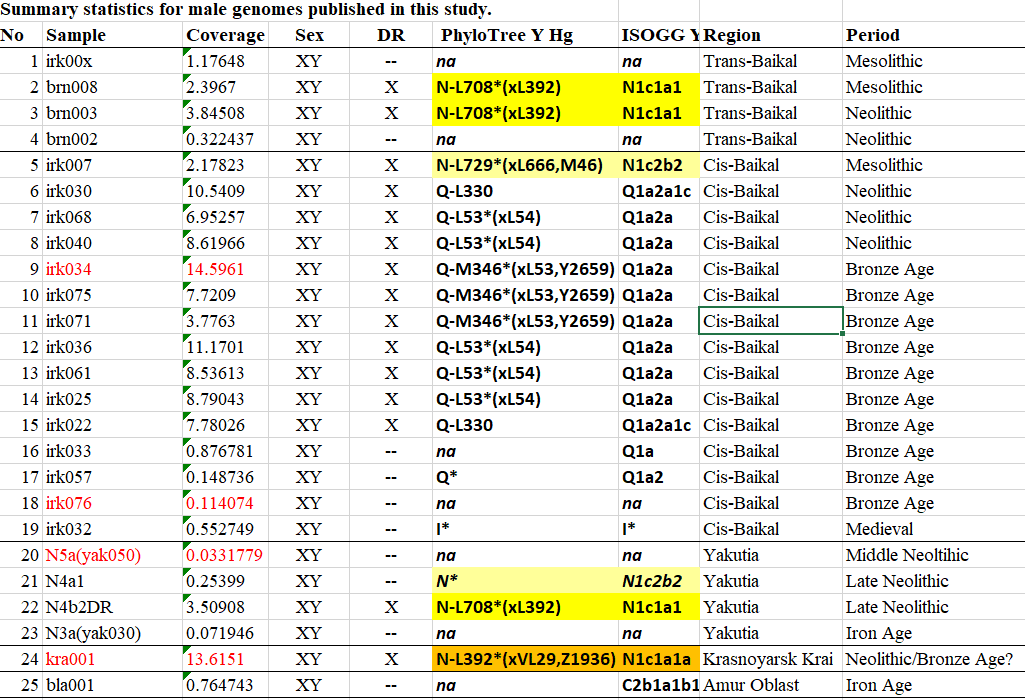
We produced whole-genome sequence data from 40 ancient individuals spanning from the Late Upper Paleolithic to the Medieval era and representing five distinct administrative regions in the Russian Federation encompassing Yakutia, Trans-Baikal, Cis-Baikal, Krasnoyarsk Krai, and Amur Oblast (…) All individuals were accredited to either Y macro-haplogroup Q or N and non-African mitochondrial macrohaplogroups of M, N, and R.
Population dynamics during and after the LGM in northeast Asia
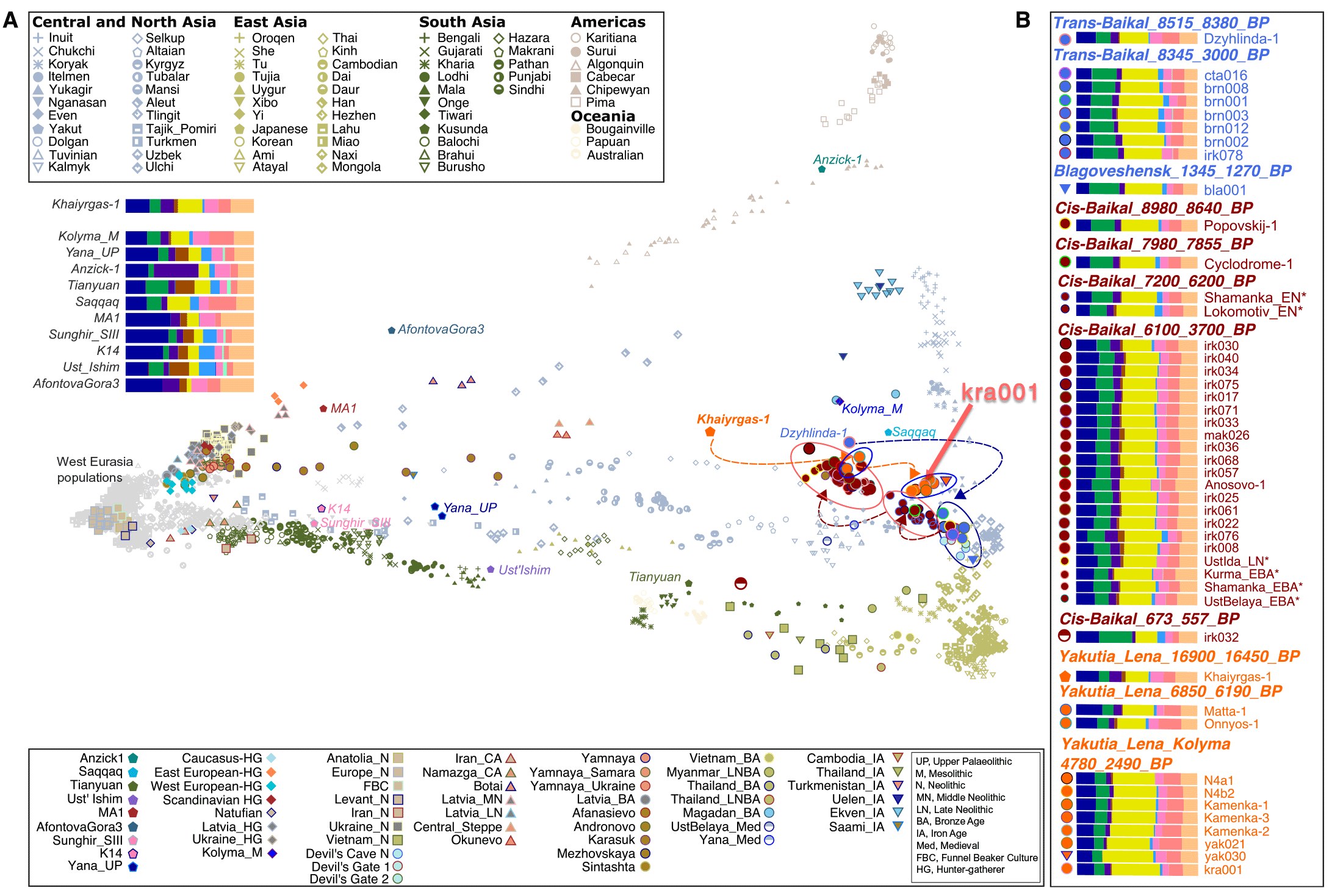
We sequenced ancient DNA from a deciduous tooth of a c.16,900-year-old subadult female (Khaiyrgas-1) excavated from the upper layer of the Paleolithic horizon of the Khaiyrgas Cave to investigate the post-LGM population dynamics in northeast Asia. This individual is one the first known post-LGM representatives of the settlers of the Central Siberian Plateau. Human groups geographically represented by this individual retracted from the area during the Bølling-Allerød warming period dated to c.15,000 to 13,000 cal BP.
Khaiyrgas-1 exhibited genetic affinity toward present-day Selkups, a north Siberian Uralic-speaking population, on the PCA. (…) On the basis of K = 14 ancestral components, a genetic component that is maximized in the present-day Nganasan population from northeast Asia (yellow) and another component that is maximized in the present-day Native America populations (purple) were present at high levels in the genome of Khaiyrgas-1 compared to other Upper Paleolithic individuals from Eurasia and Asia.
Left: Northeast Asia- and Native America-related genetic components. A subset of ADMIXTURE analysis results for K=14 ancestral clusters for Yakutia individuals compared to Native American populations (Anzick1, purple component) and Northeast Asia populations (Nganasan, yellow component). Right: ADMIXTURE result for modern populations. (Top) ADMIXTURE analysis result for K=14 ancestral clusters for present-day world populations from Human Origins data set. (Bottom) Approximate geographical positions corresponding to each of the present-day populations plotted in the fig. S4a.
Population transformations across Yakutia and the origins of the Paleo-Inuits
(…) we analyzed chronological genome sequence data from the Lena and Kolyma river regions of Yakutia. These data included a ~6800-year-old individual associated with the Syalakh cultural complex (Matta-1), a ~6200-year-old individual associated with the Belkachi cultural complex (Onnyos-1), and seven individuals dated to c.4780 to 2490 cal BP. The PCA revealed a west-east genetic cline extending from the Late Upper Paleolithic to the Iron Age and the presence of three major genetic changes during this period throughout Lena and Kolyma regions. These include a shift ~9800 years ago between the Late Upper Paleolithic and Mesolithic, another one ~6800 years ago between the Mesolithic and Early/Middle Neolithic, and ~4700 years ago between Early/Middle Neolithic and Late Neolithic/Iron Age.
Matta-1 and Onnyos-1 formed a distinct group (Yakutia_Lena_6850_6190_BP) on the PCA between Khaiyrgas-1, Kolyma_M (5), and a group of succeeding individuals (Yakutia_Lena_Kolyma_4780_2490_BP). The Bronze Age (c.4280 to 4085 BP) individual (kra001) from the Krasnoyarsk Krai grouped with Yakutia_Lena_Kolyma_4780_2490_BP. Yakutia_Lena_6850_6190_BP and Yakutia_Lena_Kolyma_4780_2490_BP exhibited genetic affinities toward present-day northeast Asia populations. ADMIXTURE analysis revealed a pattern of increase in northeast Asia–related and a pattern of decrease in Native America–related genetic ancestry from the Late Upper Paleolithic to the Iron Age. Consistently, Devil’s Cave individuals representing the East Asian ancestry and the Anzick-1 individual representing the Native American ancestry shared more alleles with the succeeding and preceding groups, respectively, implying the presence of gene flow into the Lena and Kolyma regions from Far Eastern sources during the Neolithic and the Bronze Age.
(A) PCA calculated using a set of world populations. Ancient individuals were projected onto the inferred PC space (see table S2 for information about individuals). Arrows indicate the direction of population changes in time. Asterisk denotes being published in (16). (B) A subset of ADMIXTURE result for K = 14 clusters showing the ancestral composition of investigated ancient individuals.
Origins and interactions of the Baikal populations
(…) our data revealed multiple population transformations across the Cis-Baikal area spanning a time transect between ~8980 and ~560 years ago: An ~8980-year-old Cis-Baikal individual, Popovskij-1, falls within the genetic variation of the Trans-Baikal_8345_3000_BP. Popovskij-1 shared more drift with the Trans-Baikal_8345_3000_BP compared to the other ancient groups, suggesting the presence of a uniform gene pool across the area ~8900 years ago. A ~7900-year-old individual (Cyclodrome-1) marked the first genetic shift around the Cis-Baikal region, and this individual together with individuals spanning from ~7200 to ~6200 years ago (Cis-Baikal_7200_6200_BP) formed a distinct genetic group on the PCA. Yet, Cis-Baikal individuals spanning 6100 to 3700 years ago formed another group (Cis-Baikal_6100_3700_BP), carrying comparably high amount of west Eurasia–related and low amount of northeast Asia–related components. A Medieval Cis-Baikal individual (c.670 to 550 cal BP) was genetically distinct from the other individuals with increased genetic affinities toward East Asia populations. Y haplogroup distribution was in agreement with the observed genetic groupings.
Our findings indicate a complex pattern of demographic change throughout the Baikal area consistent with (17), which revealed dynamic changes in population structure around Lake Baikal area. Before ~8500 years ago, people inhabited the Trans-Baikal region. They became ancestral to the human groups settled in Lena basin in Yakutia. About 8300 years ago, a major genetic shift happened in the Trans-Baikal area. This new group with East Asian genetic ancestry populated the west and east side of Lake Baikal during the Mesolithic. Although they remained in the Trans-Baikal region until ~3000 years ago, an extensive gene flow from distant sources affected the Cis-Baikal region ~7900 years ago and then ~6100 years ago.
Genetic diversity of ancient northeast Asia populations
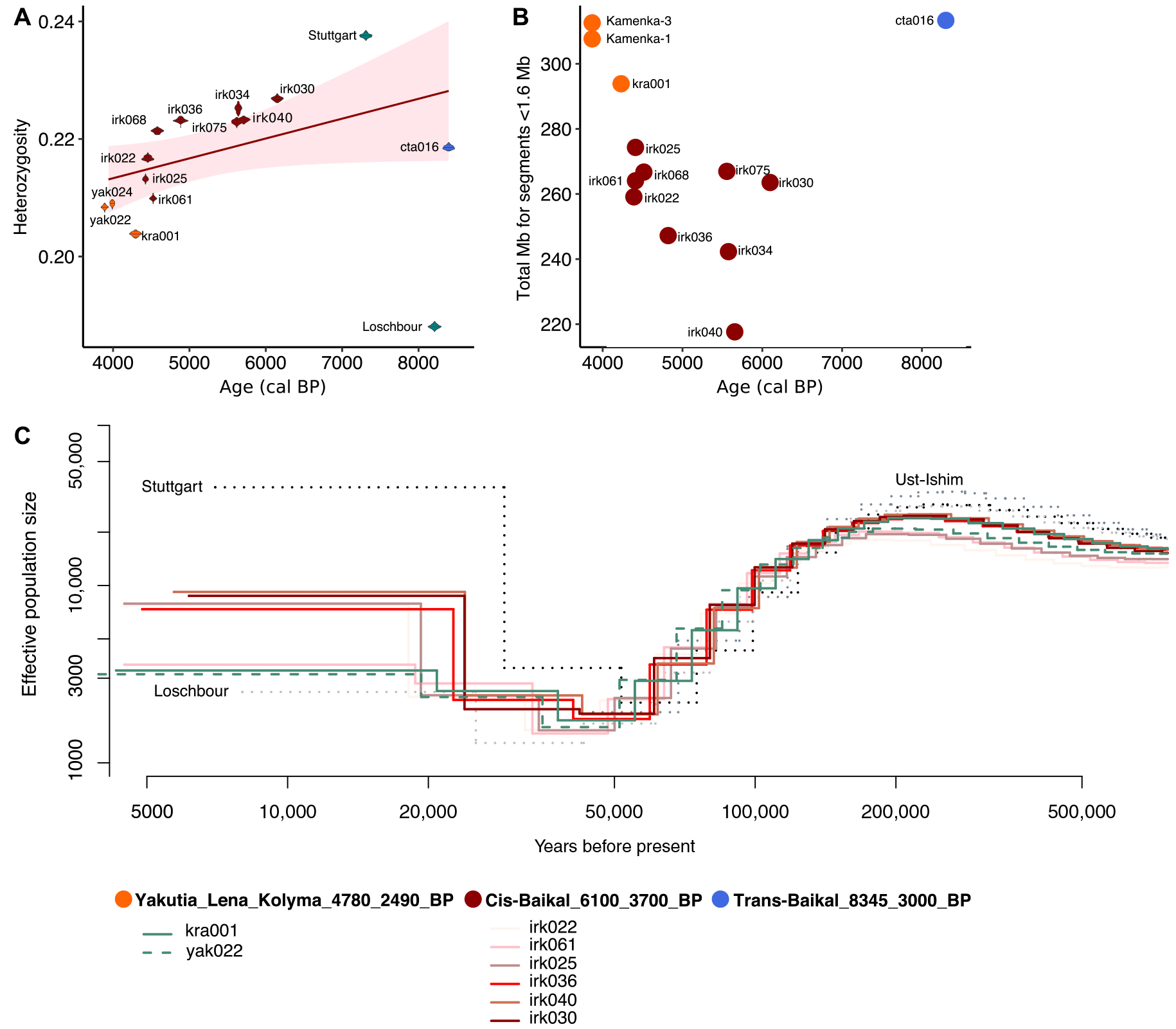
A multiple sequentially Markovian coalescent (MSMC) analysis revealed that the effective population size of Yakutia_Lena_Kolyma_4780_2490_BP was low ~4700 years ago, consistent with the low genetic diversity observed in this group. Although effective population size estimates of human groups in Cis-Baikal ~6100 to 5600 years ago were comparably high, they decreased ~4400 years ago. Similarity between the observed low effective population sizes in Cis-Baikal and Yakutia 4700 to 4400 years ago could mirror global climatic changes, i.e., cooling of the Subboreal Period, and might indicate a possible population collapse ~4700 to 4000 years ago.
Estimating the level of genetic diversity and population size changes in time. (A) Heterozygosity estimates. (B) Amount of short RoH in the genomes of Lake Baikal and Yakutia individuals. Mb, megabase. (C) Effective population size change in time estimated using MSMC.
Yersinia pestis in ancient northeast Asia
We identified 9395 Y. pestis–specific sequencing reads (3.66% genome coverage for CO92) in a ~4400-year-old individual from Cis-Baikal, Anosovo-1, and 4176 Y. pestis–specific sequencing reads (1.65% genome coverage for CO92) in a ~3800-year-old individual from Yakutia, Kamenka-2. The Kamenka burial contained three young individuals, all related to each other including a parent-child kin.
NOTE. Even though the most parsimonious interpretation – as suggested by the paper’s visual chronological data – is that these strains are related to the expansion of Afanasievo (due to the oldest Y. pestis genomes being found in Rise509 and Rise511, from Afanasievo Gora) as far as I can see the paper lacks a proper phylogenetic tree of the samples, as found e.g. in Rascovan et al. Cell (2018).
Comments
Sample kra001 (ca. 2295-2140 calBC), from Nefteprovod-2 site, burial 1, labelled Krasnoyarsk Krai LN?/EBA, is thus the earliest reported individual of hg. N-L1026*, basal (apparently) to N-VL29 and N-Z1936, while the other (basal?) N-L708* also suggest Neolithic and Bronze Age northward migration events through the Lena and Yenisei river basins likely connected to Circum-Baikal region.
NOTE. As of the time of this post, files have not been posted yet to ENA for a proper check, though. The mtDNA haplogroup for this and other samples were reported in the previous paper by Kılınç et al. Sci. Rep. (2018), and were further refined by YFull, as can be seen from the Ancient DNA Dataset.
Regarding this site from the Yenisei river basin, it seems to be formed by (at least) two different archaeological layers, one LN/EBA, the other MBA. Given the reported radiocarbon dating, burial 1 might belong to earlier, Glazkovo-related strata, immediately before the arrival of Uralic-speaking Karasuk population. A recent summary of the available literature is found in Slepchenko et al. (2017):
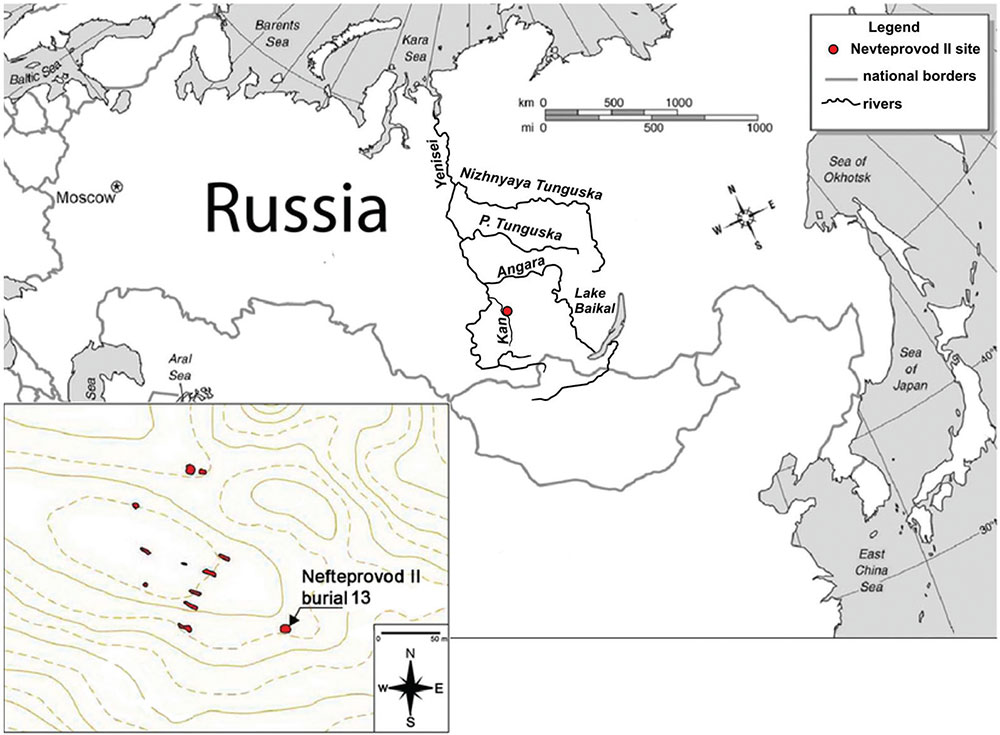
Nefteprovod I and the adjacent Nefteprovod II burial grounds were excavated as part of a salvage project at the Anzhevsk archaeological site. This site is located near the former Anzhevka settlement on the right bank of the Kan River southeast of Kansk in the Krasnoyarsk region (…) Although the recovered artefact styles have no direct analogies, similar bronze and bone artefacts and ceramic vessels are found along the middle Yenisei River and date to the Late Bronze Age. Based on these artefact similarities and the burial style, the Nefteprovod I and II burial grounds likely belong to the Karasuk culture, which succeeded the Andronovo culture in this region during the later portion of the second millennium BC. The Karasuk culture lasted from approximately 1500 to 800 BC.
Location of the archaeological site Neftprovod II. Distribution of burials on the archaeological site Neftprovod II in 2015. Burials are marked in red.
Burial #13 from the Nefteprovod II archeological site differed considerably from the other burials in its funerary style. The body was tightly flexed inside a tight burial pit with uneven margins. Grave goods were absent. The stonework overlaying the burial dated back to the Late Bronze Age, indicating that the burial came from earlier cultural strata, likely the middle Bronze Age (2000 to 1000 BC). The positioning of the body in a foetal position is unique in the region of the Kansk forest steppe. Similar burials under stonework have been found in adjacent territories of the Baikal and Angara regions, and they are associated with the Glazkovskaya culture, which dated from 2000 to 1300 BC. (Okladnikov 1975a, 1975b, Dudarёk & Lohov 2014, Derevianko et al. 2015). Tightly flexed burials were also part of the Andronovskaya culture from the Minusinsk Basin (Maksimenkov 1978).
It should come as no surprise by now to the readers of this blog that different waves of expansion of N-Tat lineages spread from around Lake Baikal. In fact, the earliest N-Tat sample known to date comes from the Trans-Baikal region, as recently reported by Ning et al. 2020 (which I referenced here).
Most known N-Tat subclades appearing in ancient samples to the west of Siberia appear therefore to be connected one way or another with westward waves of (mainly “Altaic”- or Iranian-speaking) forest-steppe- or steppe-related populations ultimately linked to the Circum-Baikal area, as explained in some of the posts listed below. In fact, the only earlier wave whose origin remained unexplained, of Palaeo-Laplandic-speaking MBA Lovozero-related BOO individuals, will probably be now easily explained as a migration linked to this early bottlenecked Cis-BaikalYakutia_Lena_Kolyma_4780_2490_BP group, to which kra001 belongs.
It is good practice to be registered
and logged in to comment.
Please keep the discussion of this post on topic. Civilized discussion. Academic tone.
For other topics, use the forums instead.