New paper (behind paywall), Genomic history of the Sardinian population, by Chiang et al. Nature Genetics (2018), previously published as a preprint at bioRxiv (2016).
#EDIT (18 Sep 2018): Link to read paper for free shared by the main author.
Interesting excerpts (emphasis mine):
Our analysis of divergence times suggests the population lineage ancestral to modern-day Sardinia was effectively isolated from the mainland European populations ~140–250 generations ago, corresponding to ~4,300–7,000 years ago assuming a generation time of 30 years and a mutation rate of 1.25 × 10−8 per basepair per generation. (…) in terms of relative values, the divergence time between Northern and Southern Europeans is much more recent than either is to Sardinia, signaling the relative isolation of Sardinia from mainland Europe.
We documented fine-scale variation in the ancient population ancestry proportions across the island. The most remote and interior areas of Sardinia—the Gennargentu massif covering the central and eastern regions, including the present-day province of Ogliastra— are thought to have been the least exposed to contact with outside populations. We found that pre-Neolithic hunter-gatherer and Neolithic farmer ancestries are enriched in this region of isolation. Under the premise that Ogliastra has been more buffered from recent immigration to the island, one interpretation of the result is that the early populations of Sardinia were an admixture of the two ancestries, rather than the pre-Neolithic ancestry arriving via later migrations from the mainland. Such admixture could have occurred principally on the island or on the mainland before the hypothesized Neolithic era influx to the island. Under the alternative premise that Ogliastra is simply a highly isolated region that has differentiated within Sardinia due to genetic drift, the result would be interpreted as genetic drift leading to a structured pattern of pre-Neolithic ancestry across the island, in an overall background of high Neolithic ancestry.
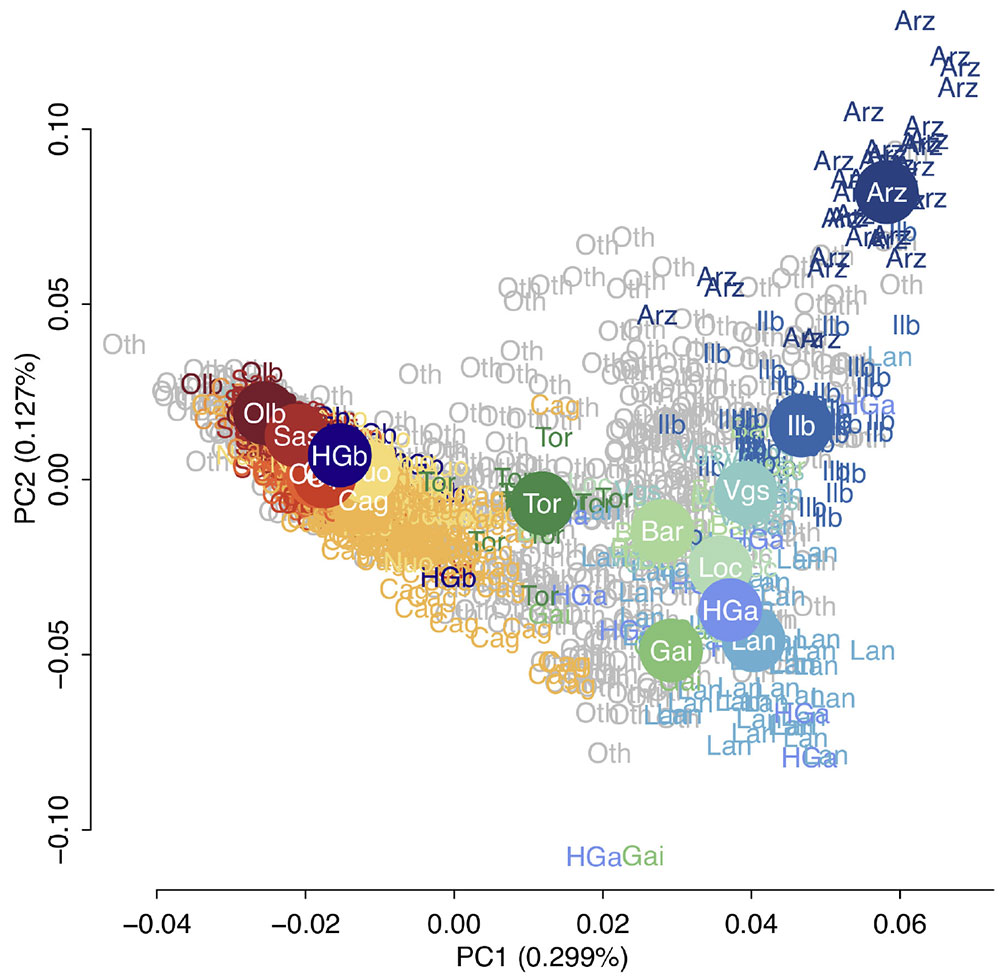
We found Sardinians show a signal of shared ancestry with the Basque in terms of the outgroup f3 shared-drift statistics. This is consistent with long-held arguments of a connection between the two populations, including claims of Basque-like, non-Indo-European words among Sardinian placenames. More recently, the Basque have been shown to be enriched for Neolithic farmer ancestry and Indo-European languages have been associated with steppe population expansions in the post-Neolithic Bronze Age. These results support a model in which Sardinians and the Basque may both retain a legacy of pre-Indo-European Neolithic ancestry. To be cautious, while it seems unlikely, we cannot exclude that the genetic similarity between the Basque and Sardinians is due to an unsampled pre-Neolithic population that has affinities with the Neolithic representatives analyzed here.
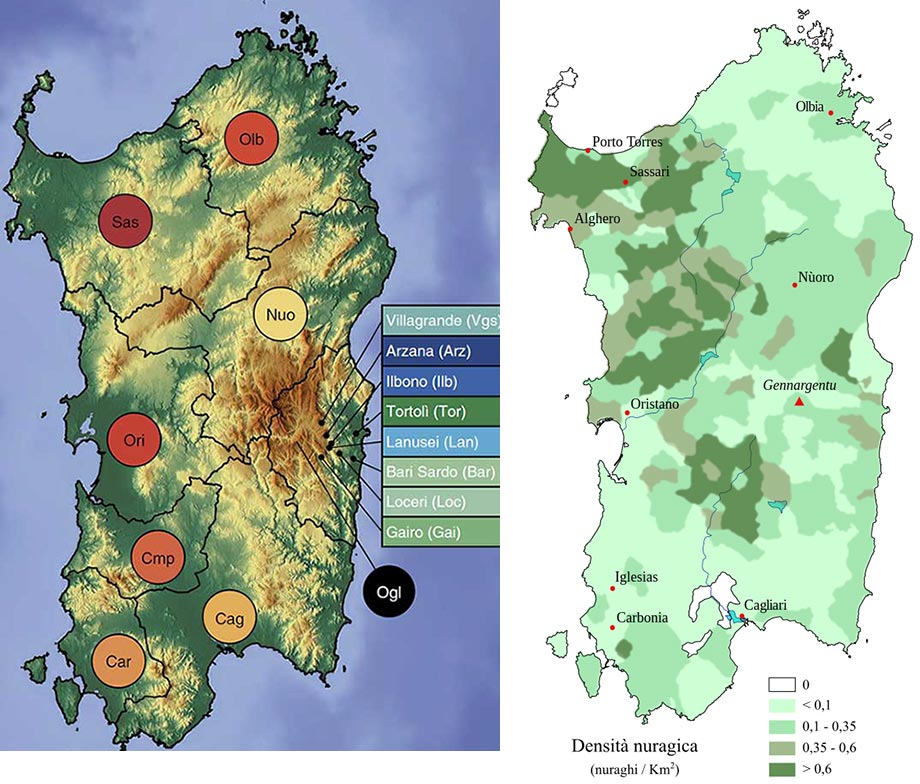
Right: Density of Nuraghi in Sardinia, from Wikipedia.
{kind=link}
While we can confirm that Sardinians principally have Neolithic ancestry on the autosomes, the high frequency of two Y-chromosome haplogroups (I2a1a1 at ~39% and R1b1a2 at ~18%) that are not typically affiliated with Neolithic ancestry is one challenge to this model. Whether these haplogroups rose in frequency due to extensive genetic drift and/or reflect sex-biased demographic processes has been an open question. Our analysis of X chromosome versus autosome diversity suggests a smaller effective size for males, which can arise due to multiple processes, including polygyny, patrilineal inheritance rules, or transmission of reproductive success. We also find that the genetic ancestry enriched in Sardinia is more prevalent on the X chromosome than the autosome, suggesting that male lineages may more rapidly trace back to the mainland. Considering that the R1b1a2 haplogroup may be associated with post-Neolithic steppe ancestry expansions in Europe, and the recent timeframe when the R1b1a2 lineages expanded in Sardinia, the patterns raise the possibility of recent male-biased steppe ancestry migration to Sardinia, as has been reported among mainland Europeans at large (though see Lazaridis and Reich and Goldberg et al.). Such a recent influx is difficult to square with the overall divergence of Sardinian populations observed here.
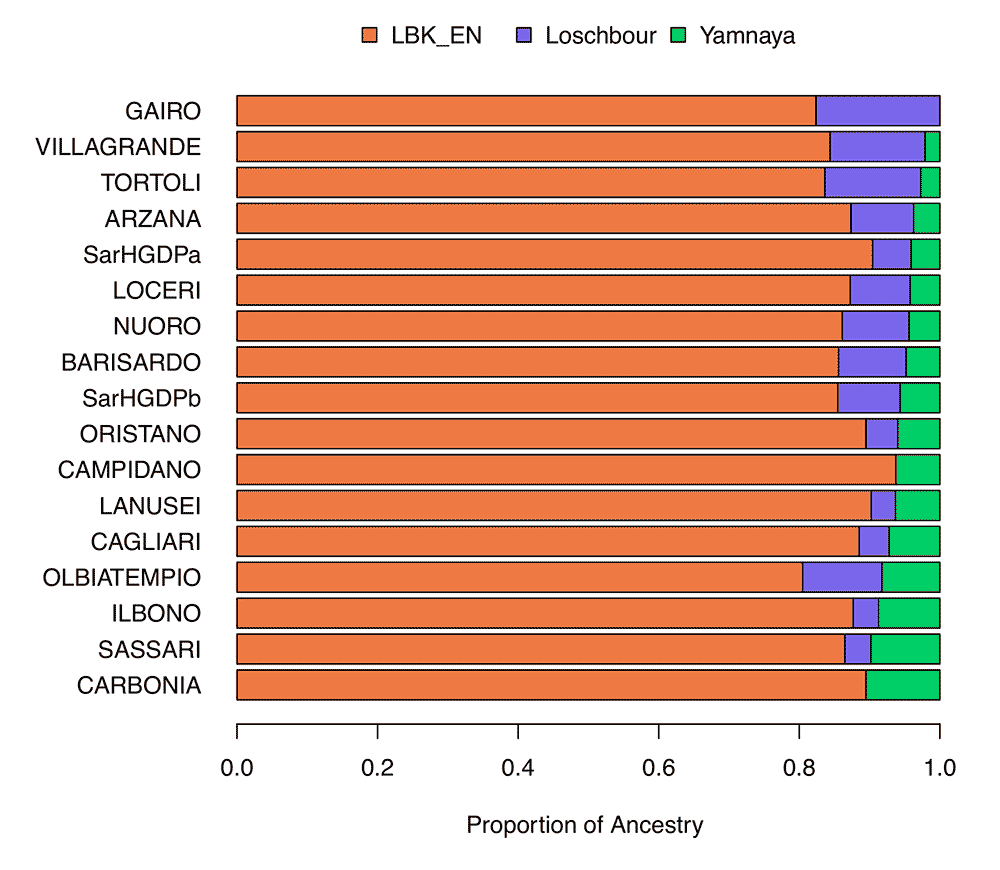
Once again, haplogroup R1b1a2 (M269), and only R1b1a2, related to male-biased, steppe-related Indo-European migrations…just sayin’.
Interestingly, haplogroup I2a1a1 is actually found among northern Iberians during the Neolithic and Chalcolithic, and is therefore associated with Neolithic ancestry in Iberia, too, and consequently – unless there is a big surprise hidden somewhere – with the ancestry found today among Basques.
NOTE. In fact, the increase in Neolithic ancestry found in south-west Ireland with expanding Bell Beakers (likely Proto-Beakers), coupled with the finding of I2a subclades in Megalithic cultures of western Europe, would support this replacement after the Cardial and Epi-Cardial expansions, which were initially associated with G2a lineages.
I am not convinced about a survival of Palaeo-Sardo after the Bell Beaker expansion, though, since there is no clear-cut cultural divide (and posterior continuity) of pre-Beaker archaeological cultures after the arrival of Bell Beakers in the island that could be identified with the survival of Neolithic languages.
We may have to wait for ancient DNA to show a potential expansion of Neolithic ancestry from the west, maybe associated with the emergence of the Nuragic civilization (potentially linked with contemporaneous Megalithic cultures in Corsica and in the Balearic Islands, and thus with an Iberian rather than a Basque stock), although this is quite speculative at this moment in linguistic, archaeological, and genetic terms.
Nevertheless, it seems that the association of a Basque-Iberian language with the Neolithic expansion from Anatolia (see Villar’s latest book on the subject) is somehow strengthened by this paper. However, it is unclear when, how, and where expanding G2a subclades were replaced by native I2 lineages.
Related
- Ancient Phoenician mtDNA from Sardinia, Lebanon reflects settlement, genetic diversity, and female mobility
- Haplogroup J spread in the Mediterranean due to Phoenician and Greek colonizations
- Migration vs. Acculturation models for Aegean Neolithic in Genetics — still depending strongly on Archaeology
- Y-chromosome mixture in the modern Corsican population shows different migration layers
- Cogotas I Bronze Age pottery emulated and expanded Bell Beaker decoration
- Iberia in the Copper and Early Bronze Age: Cultural, demographic, and environmental analysis
- First Iberian R1b-DF27 sample, probably from incoming East Bell Beakers
- Iberian prehistoric migrations in Genomics from Neolithic, Chalcolithic, and Bronze Age
- Patterns of genetic differentiation and the footprints of historical migrations in the Iberian Peninsula
- Population substructure in Iberia, highest in the north-west territory (to appear in Nature)
- Analysis of R1b-DF27 haplogroups in modern populations adds new information that contrasts with ‘steppe admixture’ results