New paper (behind paywall) Reconstruction of Y-chromosome phylogeny reveals two neolithic expansions of Tibeto-Burman populations by Wang et al. Mol Genet Genomics (2018).
Interesting excerpts:
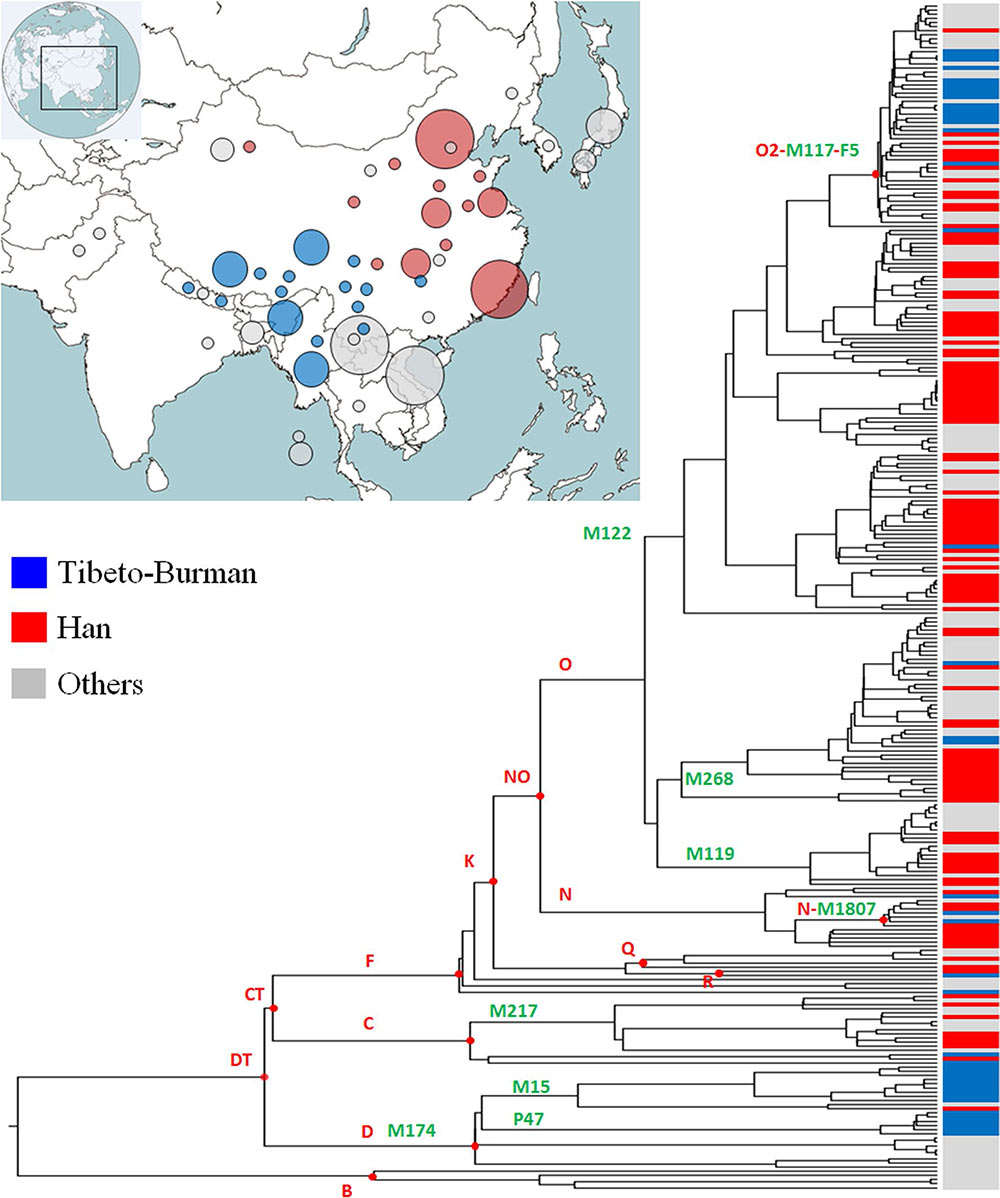
Archeological studies suggest that a subgroup of ancient populations of the Miaodigou culture (~ 6300–5500 BP) moved westward to the upper stream region of the Yellow River and created the Majiayao culture (~ 5400–4900 BP) (Liu et al. 2010), which was proposed to be the remains of direct ancestors of Tibeto-Burman populations (Sagart 2008). On the other hand, Han populations, the other major descendant group of the Yang-Shao culture (~ 7000–5500 BP), are composed of many other sub-lineages of Oα-F5 and extremely low frequencies of D-M174 (Additional files 1: Figure S1; Additional files 2: Table S1). Therefore, we propose that Oα-F5 may be one of the dominant paternal lineages in ancient populations of Yang-Shao culture and its successors.
In this study, we demonstrated that both sub-lineages of D-M174 and Oα-F5 are founding paternal lineages of modern Tibeto-Burman populations. The genetic patterns suggested that the ancestor group of modern Tibeto-Burman populations may be an admixture of two distinct ancient populations. One of them may be hunter–gatherer populations who survived on the plateau since the Paleolithic Age, represented by varied sub-lineages of sub-lineages of D-M174. The other one was comprised of farmers who migrated from the middle Yellow River basin, represented by sub-lineages of Oα-F5. In general, the genetic evidence in this study supports the conclusion that the appearance of the ancestor group of Tibeto-Burman populations was triggered by the Neolithic expansion from the upper-middle Yellow River basin and admixture with local populations on the Tibetan Plateau (Su et al. 2000).
Two neolithic expansion origins of Tibeto‑Burman populations
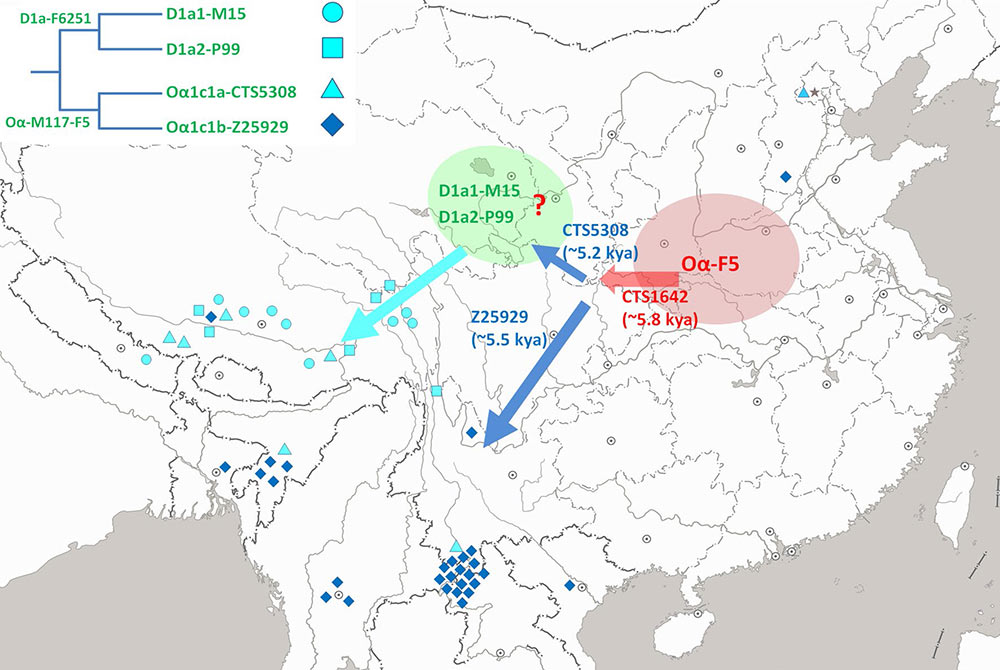
We also observed significant differences in the paternal gene pool of different subgroups of Tibeto-Burman populations. Haplogroup D-M174 contributed ~ 54% percent in a sampling of 2354 Tibetan males throughout the Tibetan Plateau (Qi et al. 2013). Previous studies have also found high frequencies of D-M174 in other populations on the Tibetan Plateau (Shi et al. 2008), including Sherpa (Lu et al. 2016) and Qiang (Wang et al. 2014). In contrast, haplogroup D-M174 is rare or absent from Tibeto-Burman populations from Northeast India and Burma (Shi et al. 2008). In populations of the Ngwi-Burmese language subgroup, the average frequencies of haplogroup D-M174 are ~ 5% (Dong et al. 2004; Peng et al. 2014). Furthermore, we found that lineage Oα1c1b-CTS5308 is mainly found in Tibeto-Burman populations from the Tibetan Plateau. In contrast, lineage Oα1c1a-Z25929 was found in Tibeto-Burman populations from Northeast India, Burma, and the Yunan and Hunan provinces of China (Additional files 1: Figure S1; Additional files 2: Table S1). In general, enrichment of lineage Oα1c1b- CTS5308 and high frequencies of D-M174 can be found in most Tibeto-Burman populations on the Tibetan Plateau and adjacent regions, whereas Tibeto-Burman populations from other regions tend to have lineage Oα1c1a-Z25929 and a little to no percentage of D-M174.
The inconsistent pattern we observed in the paternal gene pool of modern Tibeto-Burman populations suggested that there may be two distinct ancestor groups (Fig. 3). The proposed migration routes shown in Fig. 3 are somewhat different from those proposed by Su et al. (2000). According to our age estimation, most of the D1a2a-P47 samples belong to sub-lineage PH116, a young lineage that emerged ~ 2500 years ago (95% CI 1915–3188 years). On the other hand, continuous differentiation can be observed on a phylogenetic tree of lineages D1a1a1a1-PH4979 and D1a1a1a2-Z31591 since 6000 years ago. Therefore, we proposed that a group of ancient populations may have moved to the upper basin of the Yellow River and admixed intensively with local populations with high frequencies of haplogroup D-M174, including its sub-lineage D1a2a-P47 (Fig. 3). This ancestor group eventually gave birth to modern Tibeto-Burman populations on the Tibetan Plateau and adjacent regions. The other ancestor group moved toward the southwest and finally reached South East Asia (Burma and other locations) and the northeastern part of India (Fig. 3). This ancestor group may have had no or a minor admixture of D-M174 in their paternal gene pool.
Long‑term admixture before expansion to a high‑altitude region
It is interesting to investigate the time gap between the appearance of Neolithic cultures in the northeastern part of the Tibetan Plateau and the final phase of human expansion across the Tibetan Plateau. The Majiayao culture (~ 5400–4900 BP) is the earliest Neolithic culture in the northeastern part of the Tibetan Plateau (Liu et al. 2010). However, previous archeological study has suggested that the final phase of diffusion into the high-altitude area of the Tibetan Plateau occurred at approximately 3.6 kya (Chen et al. 2015). Our genetic evidence in this study is consistent with this scenario based on archeological evidence. Based on Y-chromosome analysis in this study, many unique lineages of Tibeto-Burman populations emerged between 6000 years ago and 2500 years ago (Additional files 3: Table S2). The most recent common age of D1a2-PH116, a sub-lineage that spread throughout the Tibetan Plateau, is only 2500 years ago.
We propose that there may be two important factors for the observed age gap. First, living in a high-altitude environment may require some crucial physical characteristics that were lacking from Neolithic immigrants from the middle Yellow River Basin. Intense genetic admixture with local people who had survived on the Tibetan Plateau since the Paleolithic Age may have actually guaranteed the expansion of humans across the Tibetan Plateau. Therefore, a long period of admixture, lasting from 5.4 to 3.6 kya, may be necessary for the appearance of a population with beneficial genetic variants that was genetically adapted to the high-altitude environment. Second, technological innovations, such as the domestication of wheat and highland barley (Chen et al. 2015), establishment of yak pastoralism (Rhode et al. 2007), and introduction of other culture elements in the Bronze Age (Ma et al. 2016), are also important factors that facilitated permanent settlements with large population sizes in the high-altitude area of the Tibetan Plateau.
Related:
- Bantu distinguished from Khoe by uniparental markers, not genome-wide autosomal admixture
- Post-Neolithic Y-chromosome bottleneck explained by cultural hitchhiking and competition between patrilineal clans
- Mitogenomes from Thailand offer insights into maternal genetic history of mainland South-East Asia
- Ancient genomes document multiple waves of migration in south-east Asian prehistory
- Genomics reveals four prehistoric migration waves into South-East Asia
- Population turnover in Remote Oceania shortly after initial settlement
- Language continuity despite population replacement in Remote Oceania
- Genomic history of South-East Asia: eastern Polynesians, Peninsular Malaysia and North Borneo
- Islands across the Indonesian archipelago show complex patterns of admixture
- Two more studies on the genetic history of East Asia: Han Chinese and Thailand
- Reconstructing the demographic history of the Himalayan and adjoining populations
- Ancient Di-Qiang people show early links with Han Chinese