Open access Demographic history and genetic adaptation in the Himalayan region inferred from genome-wide SNP genotypes of 49 populations, by Arciero et al. Mol. Biol. Evol (2018), accepted manuscript (msy094).
Abstract (emphasis mine):
We genotyped 738 individuals belonging to 49 populations from Nepal, Bhutan, North India or Tibet at over 500,000 SNPs, and analysed the genotypes in the context of available worldwide population data in order to investigate the demographic history of the region and the genetic adaptations to the harsh environment. The Himalayan populations resembled other South and East Asians, but in addition displayed their own specific ancestral component and showed strong population structure and genetic drift. We also found evidence for multiple admixture events involving Himalayan populations and South/East Asians between 200 and 2,000 years ago. In comparisons with available ancient genomes, the Himalayans, like other East and South Asian populations, showed similar genetic affinity to Eurasian hunter-gatherers (a 24,000-year-old Upper Palaeolithic Siberian), and the related Bronze Age Yamnaya. The high-altitude Himalayan populations all shared a specific ancestral component, suggesting that genetic adaptation to life at high altitude originated only once in this region and subsequently spread. Combining four approaches to identifying specific positively-selected loci, we confirmed that the strongest signals of high-altitude adaptation were located near the Endothelial PAS domain-containing protein 1 (EPAS1) and Egl-9 Family Hypoxia Inducible Factor 1 (EGLN1) loci, and discovered eight additional robust signals of high-altitude adaptation, five of which have strong biological functional links to such adaptation. In conclusion, the demographic history of Himalayan populations is complex, with strong local differentiation, reflecting both genetic and cultural factors; these populations also display evidence of multiple genetic adaptations to high-altitude environments.
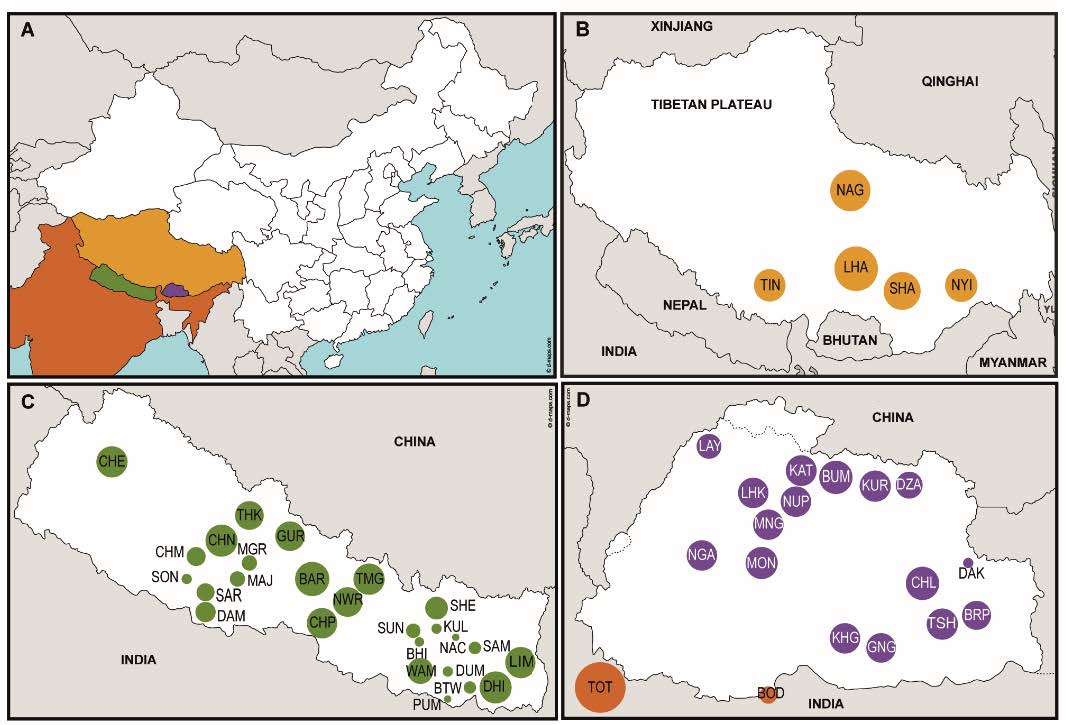
Relevant excerpts:
Genetic affinity to ancestral populations
We explored the genetic affinity between the Himalayan populations and five ancient genomes using f3-outgroup statistics. Himalayans show greater affinity to Eurasian hunter-gatherers (MA-1, a 24,000- year-old Upper Palaeolithic Siberian), and the related Bronze Age Yamnaya, than to European farmers (5,500-4,800 years ago; Fig. 5A) or to European hunter-gatherers (La Braña, 7,000 years ago; Fig. 5B), like other South and East Asian populations. We further explored the affinity of Himalayan populations by comparing them with the 45,000-year-old Upper Palaeolithic hunter-gatherer (Ust’-Ishim) and each of MA-1, La Braña, or Yamnaya. Himalayan individuals cluster together with other East Asian populations and show equal distance from Ust’-Ishim and the other ancient genomes, probably because Ust’-Ishim belongs to a much earlier period of time (supplementary fig. S15). We also explored genetic affinity between modern Himalayan populations and five ancient Himalayans (3,150 1,250 years old) from Nepal. The ancient individuals cluster together with modern Himalayan populations in a worldwide PCA (supplementary fig. S16), and the f3-outgroup statistics show modern high-altitude populations have the closest affinity with these ancient Himalayans, suggesting that these ancient individuals could represent a proxy for the first populations residing in the region (supplementary fig. S17 and supplementary table S4). Finally, we explored the genetic affinity of Himalayan samples with the archaic genomes of Denisovans and Neanderthals (Skoglund and Jakobsson 2011), and found that they show a similar sharing pattern with Denisovans and Neanderthals to the other South and East Asian populations. Individuals belonging to four Nepalese, one Cambodian, and three Chinese populations show the highest Denisovan sharing (after populations from Australia and Papua New Guinea) but these values are not significantly greater than other South and East Asian populations (supplementary figs. S18 and S19).
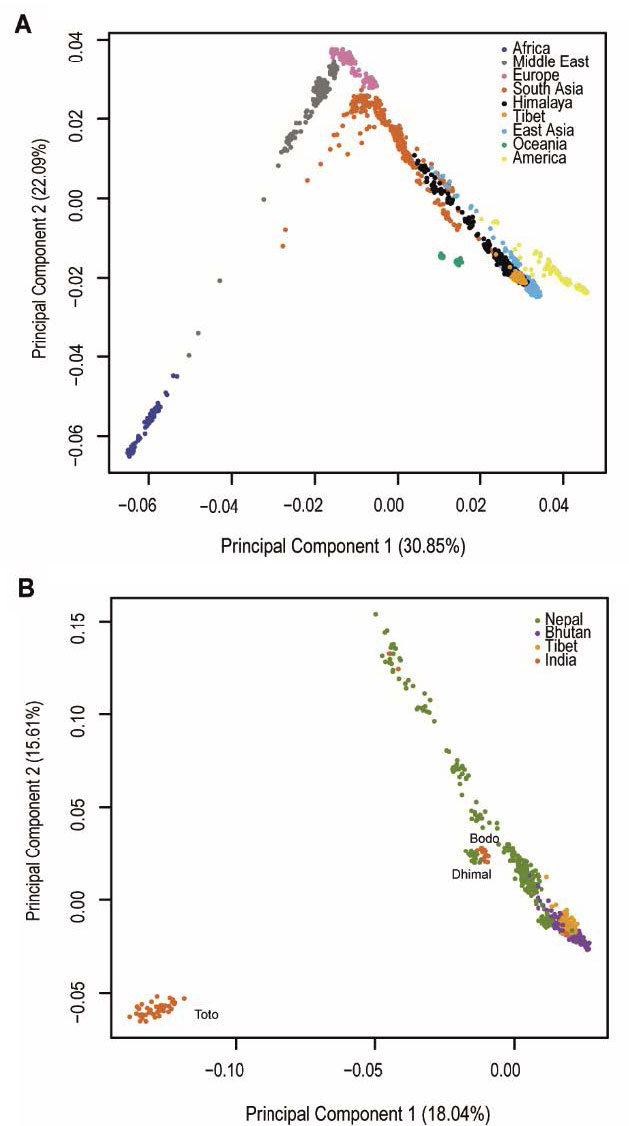
NOTE. The variance explained in the PCA graphics seems to be too high. This happened recently also with the Damgaard et al. (2018) papers (see here the comment by Iosif Lazaridis).
Similarities and differences between high-altitude Himalayan
The most striking example is provided by the Toto from North India, an isolated tribal group with the lowest genetic diversity of the Himalayan populations examined here, indicated by the smallest long-term Ne (supplementary fig. S5), and a reported census size of 321 in 1951 (Mitra 1951), although their numbers have subsequently increased. Despite this extreme substructure, shared common ancestry among the high-altitude populations (Fig. 2C and Fig. 3) can be detected, and the Nepalese in general are distinguished from the Bhutanese and Tibetans (Fig. 2C) and they also cluster separately (Fig. 3). In a worldwide context, they share an ancestral component with South Asians (supplementary fig. S2). On the other hand, the Tibetans do not show detectable population substructure, probably due to a much more recent split in comparison with the other populations (Fig. 2C and supplementary fig. S6). The genetic similarity between the high-altitude populations, including Tibetans, Sherpa and Bhutanese, is also supported by their clustering together on the phylogenetic tree, the PCA generated from the co-ancestry matrix generated by fineSTRUCTURE (supplementary fig. S10 and S11), the lack of statistical significance for most of the D-statistics tests (Yoruba, Han; high-altitude Himalayan 1, high-altitude Himalayan 2), and the absence of correlation between the increased genetic affinity to lowland East Asians and the spatial location of the Himalayan populations (supplementary figs. S12 and S13). Together, these results suggest the presence of a single ancestral population carrying advantageous variants for high-altitude adaptation that separated from lowland East Asians, and then spread and diverged into different populations across the Himalayan region. (…)
Recent admixture events
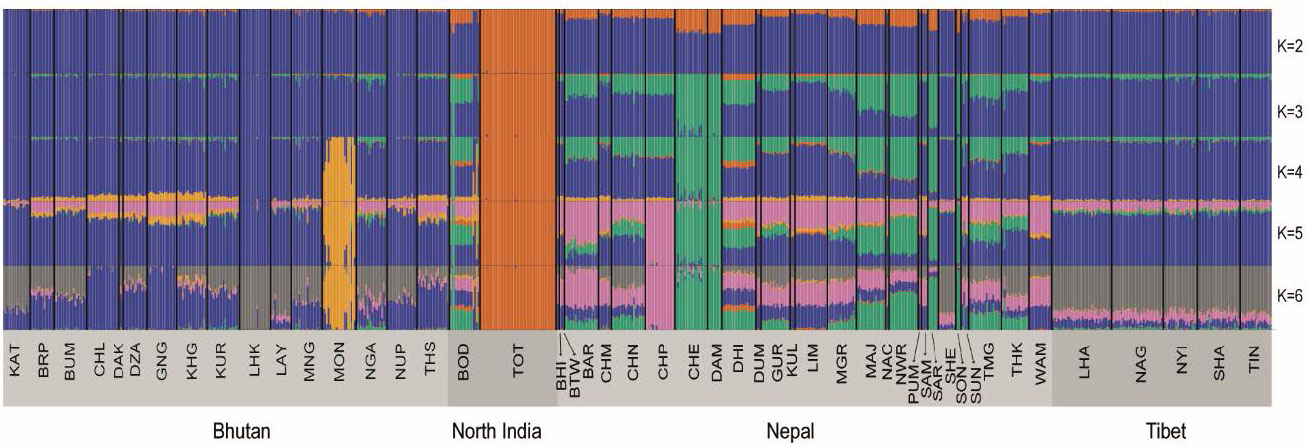
Himalayan populations show signatures of recent admixture events, mainly with South and East Asian populations as well as within the Himalayan region itself. Newar and Lhasa show the oldest signature of admixture, dated to between 2,000 and 1,000 years ago. Majhi and Dhimal display signatures of admixture within the last 1,000 years. Chetri and Bodo show the most recent admixture events, between 500 and 200 years ago (Fig. 4, supplementary tables S3). The comparison between the genetic tree and the linguistic association of each Himalayan population highlights the agreement between genetic and linguistic sub-divisions, in particular in the Bhutanese and Tibetan populations. Nepalese populations show more variability, with genetic sub-clusters of populations belonging to different linguistic affiliations (Fig. 3B). Modern high-altitude Himalayans show genetic affinity with ancient genomes from the same region (supplementary fig. S17), providing additional support for the idea of an ancient high-altitude population that spread across the Himalayan region and subsequently diverged into several of the present-day populations. Furthermore, Himalayan populations show a similar pattern of allele sharing with Denisovans as other South-East Asian populations (supplementary fig. S18 and S19). Overall, geographical isolation, genetic drift, admixture with neighbouring populations and linguistic subdivision played important roles in shaping the genetic variability we see in the Himalayan region today.
Related:
- Reconstructing the demographic history of the Himalayan and adjoining populations
- No large-scale steppe migration into Anatolia; early Yamna migrations and MLBA brought LPIE dialects in Asia
- Eurasian steppe dominated by Iranian peoples, Indo-Iranian expanded from East Yamna
- Early Indo-Iranian formed mainly by R1b-Z2103 and R1a-Z93, Corded Ware out of Late PIE-speaking migrations
- Mitogenomes show ancient human migrations to and through North-East India not of males exclusively
- The Indus Valley Civilisation in genetics – the Harappan Rakhigarhi project
- Genomics reveals four prehistoric migration waves into South-East Asia
- The Aryan migration debate, the Out of India models, and the modern “indigenous Indo-Aryan” sectarianism
- Indo-European and Central Asian admixture in Indian population, dependent on ethnolinguistic and geodemographic divisions
- Genomic history of South-East Asia: eastern Polynesians, Peninsular Malaysia and North Borneo
- Ancient genomes document multiple waves of migration in south-east Asian prehistory