Preprint Late Pleistocene human genome suggests a local origin for the first farmers of central Anatolia, by Feldman et al. bioRxiv (2018).
Interesting excerpts (emphasis mine):
Anatolian hunter-gatherers experienced climatic changes during the last glaciation and inhabited a region that connects Europe to the Near East. However, interactions between Anatolia and Southeastern Europe in the later Upper Palaeolithic/Epipalaeolithic are so far not well documented archaeologically. Interestingly, a previous genomic study showed that present-day Near-Easterners share more alleles with European hunter-gatherers younger than 14,000 BP (‘Later European HG’) than with earlier ones (‘Earlier European HG’). With ancient genomic data available, we could directly compare the Near-Eastern hunter-gatherers (AHG and Natufian) with the European ones. As is the case for present-day Near-Easterners, the Near-Eastern hunter-gatherers share more alleles with the Later European HG than with the Earlier European HG, shown by the significantly positive statistic D(Later European HG, Earlier European HG; AHG/Natufian, Mbuti). Among the Later European HG, recently reported Mesolithic hunter-gatherers from the Balkan peninsula, which geographically connects Anatolia and central Europe (‘Iron Gates HG’), are genetically closer to AHG when compared to all the other European hunter-gatherers, as shown in the significantly positive statistic D(Iron_Gates_HG, European hunter-gatherers; AHG, Mbuti/Altai). Iron Gates HG are followed by Epigravettian and Mesolithic individuals from Italy and France (Villabruna and Ranchot respectively) as the next two European hunter-gatherers genetically closest to AHG. Iron Gates HG have been suggested to be genetically intermediate between WHG and eastern European hunter-gatherers (EHG) with an additional unknown ancestral component.
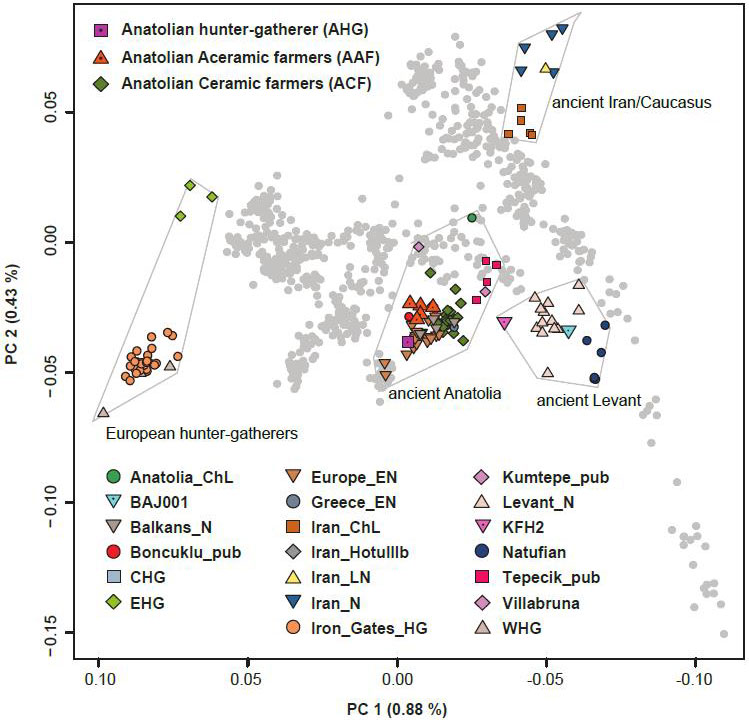
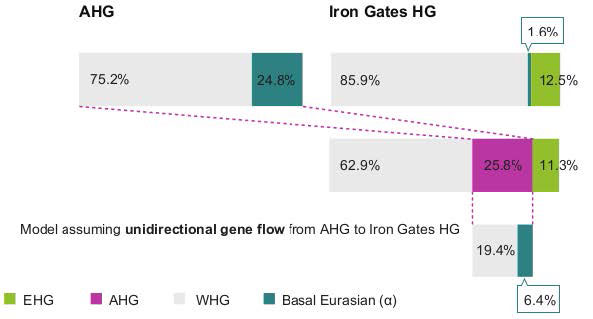
We find that Iron Gates HG can be modeled as a three-way mixture of Near-Eastern hunter-gatherers (25.8 ± 5.0 % AHG or 11.1 ± 2.2 % Natufian), WHG (62.9 ± 7.4 % or 78.0 ± 4.6 % respectively) and EHG (11.3 ± 3.3 % or 10.9 ± 3 % respectively). The affinity detected by the above D-statistic can be explained by gene flow from Near-Eastern hunter-gatherers into the ancestors of Iron Gates or by a gene flow from a population ancestral to Iron Gates into the Near-Eastern hunter-gatherers as well as by a combination of both. To distinguish the direction of the gene flow, we examined the Basal Eurasian ancestry 5 component (α), which is prevalent in the Near East but undetectable in European hunter-gatherers. Following a published approach, we estimated α to be 24.8 ± 5.5 % in AHG and 38.5 ± 5.0 % in Natufians, consistent with previous estimates for the latter. Under the model of unidirectional gene flow from Anatolia to Europe, 6.4 % is expected for α of Iron Gates by calculating (% AHG in Iron Gates HG) × (α in AHG). However, Iron Gates can be modeled without any Basal Eurasian ancestry or with a non-significant proportion of 1.6 ± 2.8 %, suggesting that unidirectional gene flow from the Near East to Europe alone is insufficient to explain the extra affinity between the Iron Gates HG and the Near-Eastern hunter-gatherers. Thus, it is plausible to assume that prior to 15,000 years ago there was either a bidirectional gene flow between populations ancestral to Southeastern Europeans of the early Holocene and Anatolians of the late glacial or a dispersal of Southeastern Europeans into the Near East. Presumably, this Southeastern European ancestral population later spread into central Europe during the post-last-glacial maximum (LGM) period, resulting in the observed late Pleistocene genetic affinity between the Near East and Europe.
While ancestry is not always relevant to distinguish certain population movements (see here), especially – as in this case – when there are few samples (thus neither geographically nor chronologically representative) and no previous model to test, it seems that ancestry and Y-DNA show a great degree of continuity in Anatolia since the Palaeolithic until the Neolithic, at least in the sampled regions. C1a2 appears in Europe since ca. 40,000 years ago (viz. Kostenki, Goyet, Vestonice, etc., and later emerges again in the Balkans after the Anatolian Neolithic expansion, probably a resurge of European groups).
The potential transition of a G2a-dominated agricultural society – that is later prevalent in Anatolian and European farmers – may have therefore happened during the Aceramic III period (ca. 8000 BC), a process of haplogroup expansion probably continuing through the early part of the Pottery Neolithic, as the society based on kinship appeared (Rosenberg and Erim-Özdoğan 2011). There is still much to know about the spread of ceramic technology and southwestern Asia domesticate complex, though.
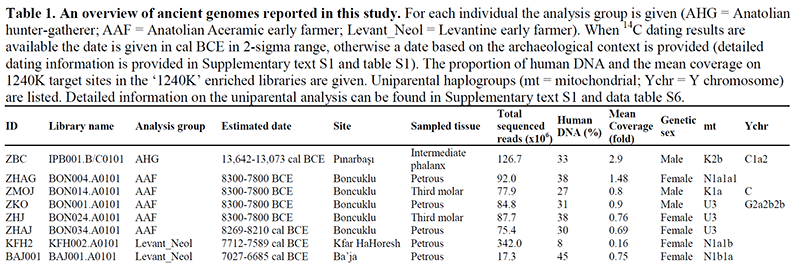
Without a proper geographical sampling, representative of previous and posterior populations, it is impossible to say. But the expansion of R1b-L754 through Anatolia to form part of the Villabruna cluster (and also the Iron Gates HG) seems perfectly possible with this data, although this paper does not help clarify the when or how. We have seen significant changes in ancestry happen within centuries with expanding populations admixing with locals. Palaeolithic sampling – like this one – shows few individuals scattered geographically over thousands of km and chronologically over thousands of years…
Related
- Agricultural origins on the Anatolian plateau
- Palaeolithic and Mesolithic connections between Italy and the Balkans, and the potential expansion of the Villabruna cluster and haplogroup R1b in Europe
- Sahara’s rather pale-green and discontinuous Sahelo-Sudanian steppe corridor, and the R1b – Afroasiatic connection
- The Danube Corridor Hypothesis and the Carpathian Basin in the Aurignacian
- Migration vs. Acculturation models for Aegean Neolithic in Genetics — still depending strongly on Archaeology
- Consequences of Damgaard et al. 2018 (I): EHG ancestry in Maykop samples, and the potential Anatolian expansion routes