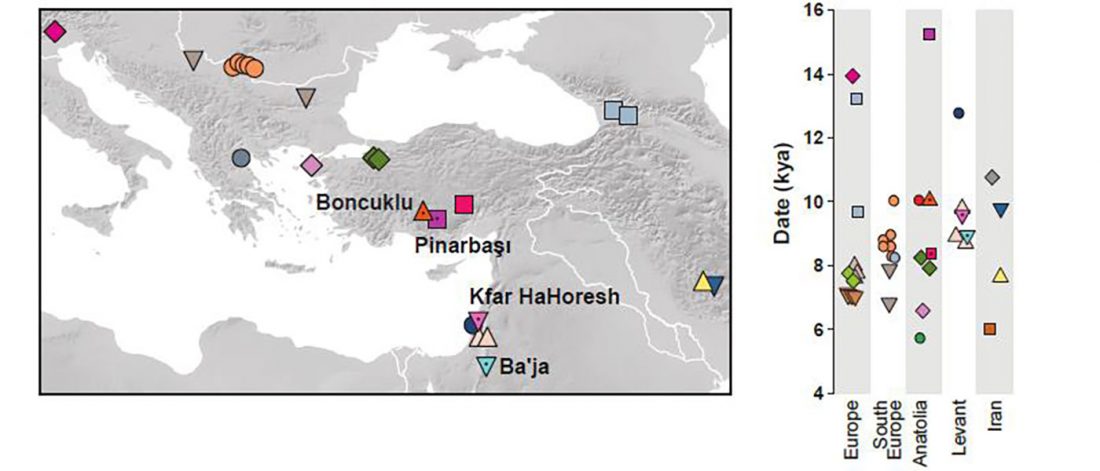
This post is part of a draft on palaeolinguistics and the Proto-Uralic homeland. See below for the color code of protoforms.
1. Foraging
1.1. Hunting
PFU (Fi., Ma., Md., Ms., Hu.) *lewi- ‘to shoot’ (UEW Nº 482; Aikio 2019: 47).
PU (Saa., Fi., Md., Ma., Kh., Ms., Hu., Smy.) *je̮ŋsi/*joŋsi ‘bow’ (UEW Nº 190; Aikio 2020: 71 with references).
PU (Saa.? Fi., Ma., Kh., Ms., Hu., Smy.) *jänti(ni) ‘sinew; (bow)string’ (UEW Nº 169; Aikio 2019: 47 with references).
PU (Saa., Fi., Md.., Ma., P, Kh., Ms., Hu., Smy.) *ńe̮li (*ńōle) ‘arrow’ (UEW Nº 622).… Read the rest “Proto-Uralic Homeland (I): Foraging & Animals”