New paper (behind paywall) Ancient Rome: A genetic crossroads of Europe and the Mediterranean, by Antonio et al. Science (2019).
The paper offers a lot of interesting data concerning the Roman Empire and more recent periods, but I will focus on Italic and Etruscan origins.
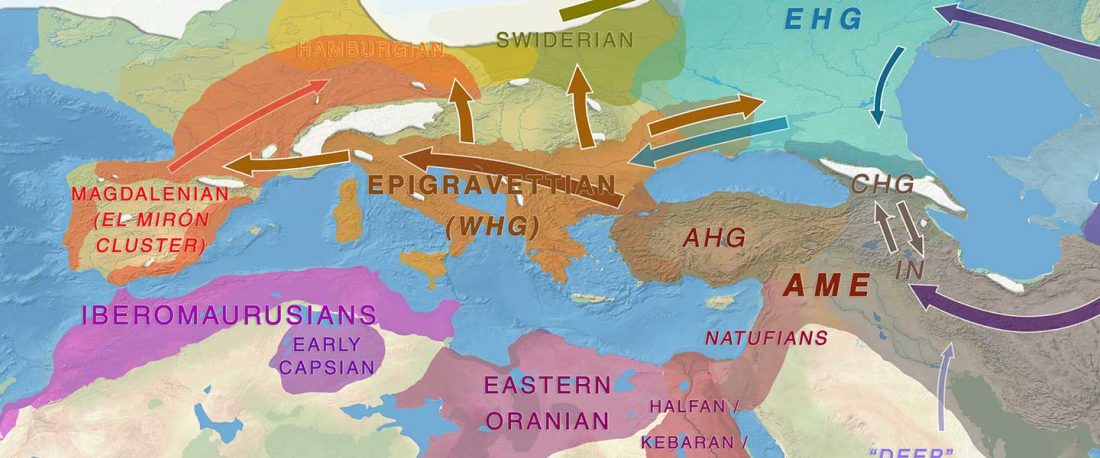
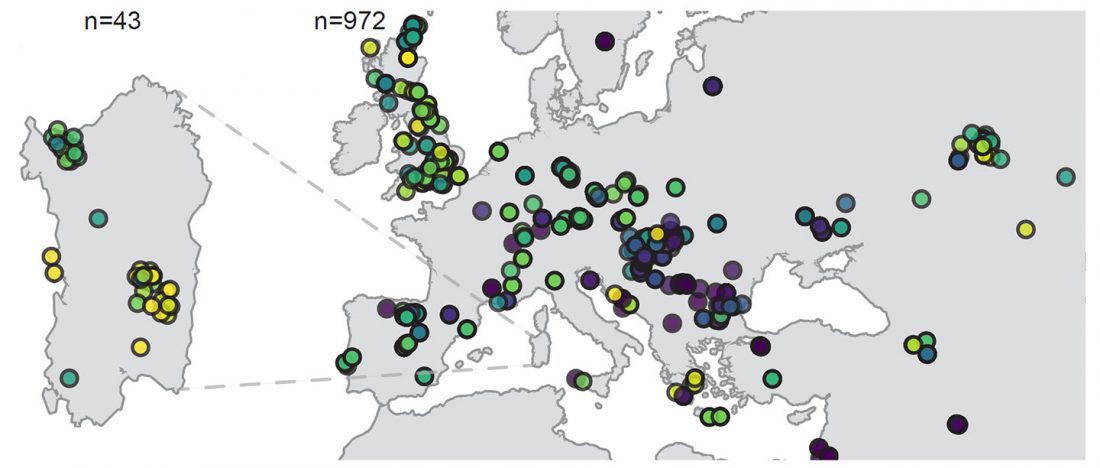
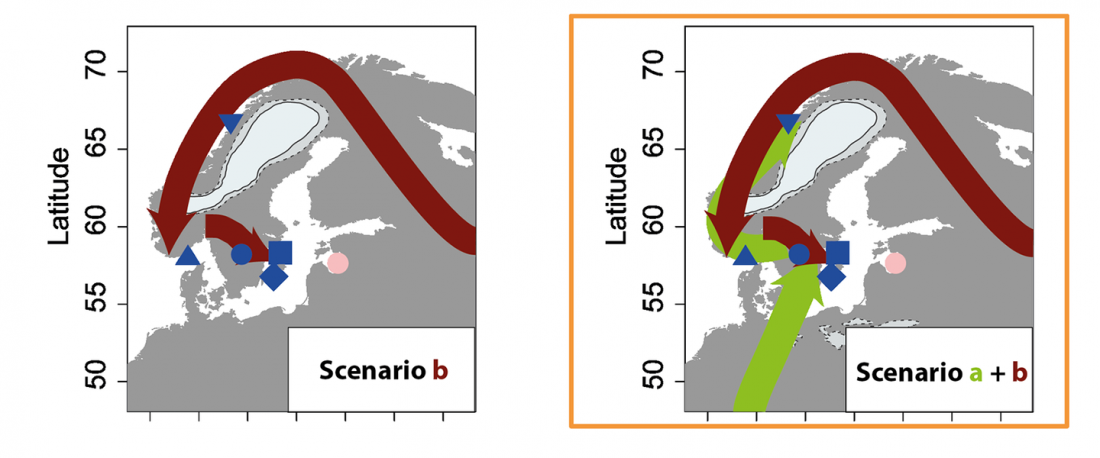
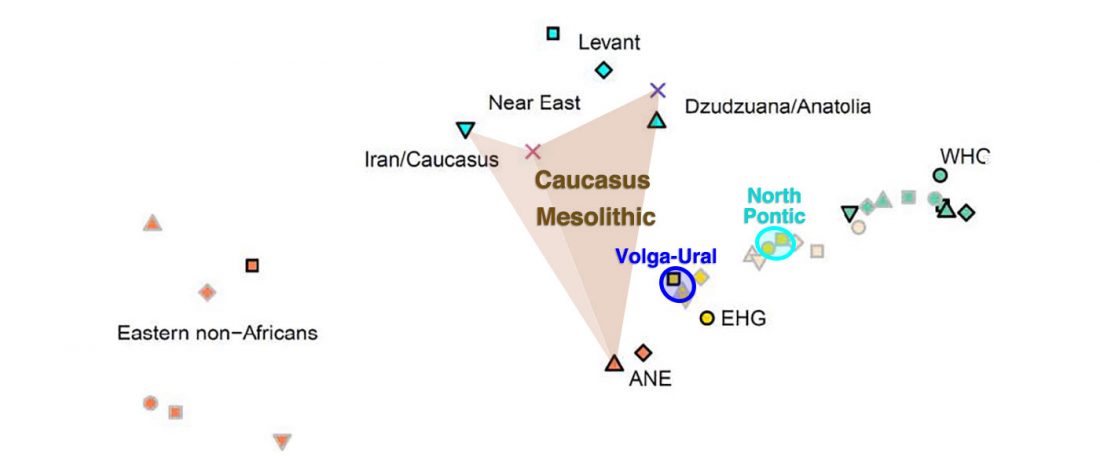
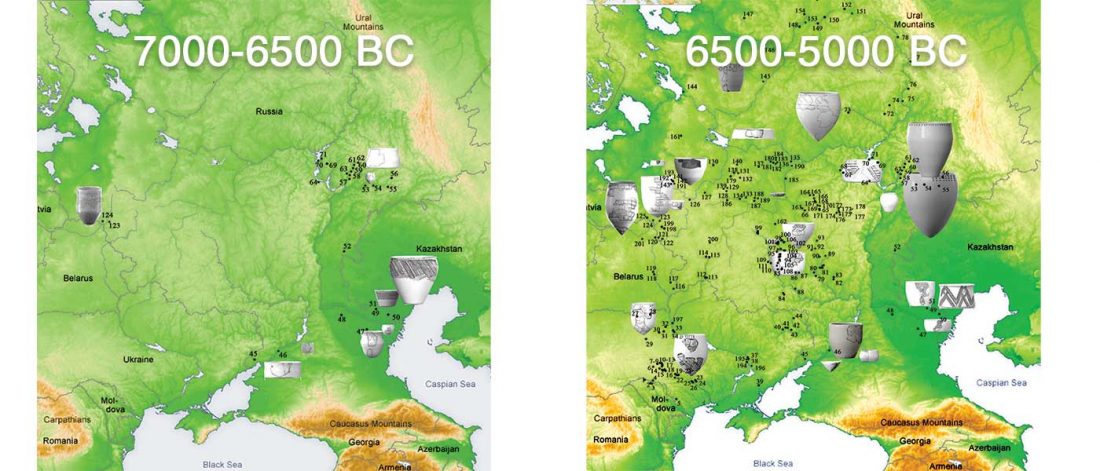
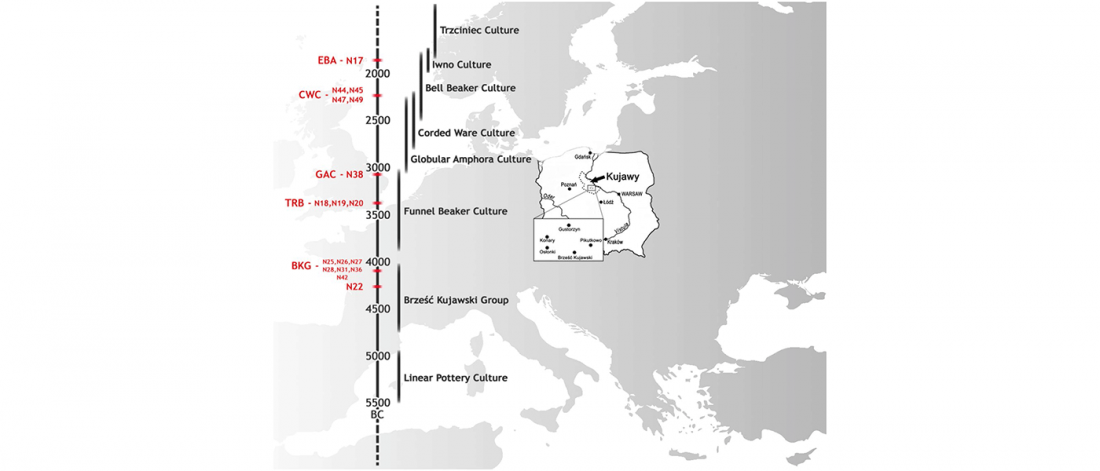
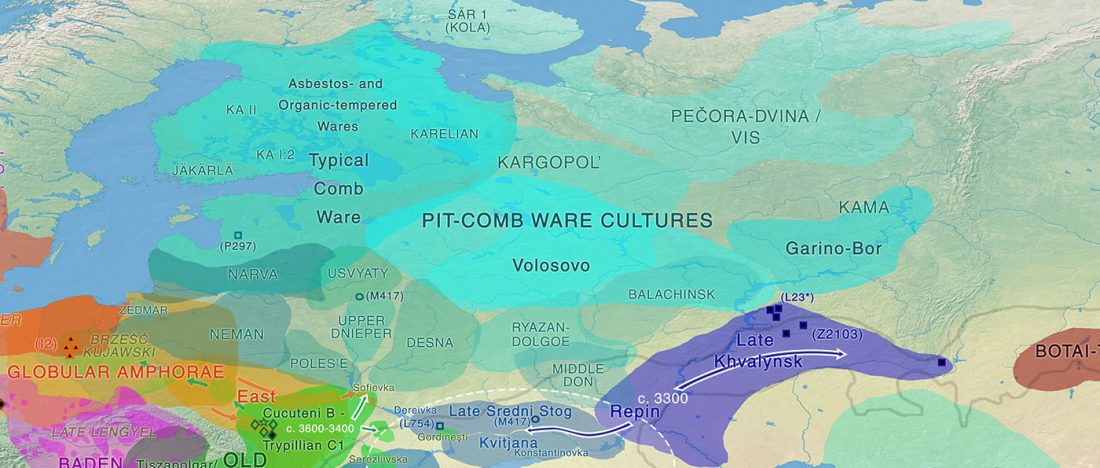
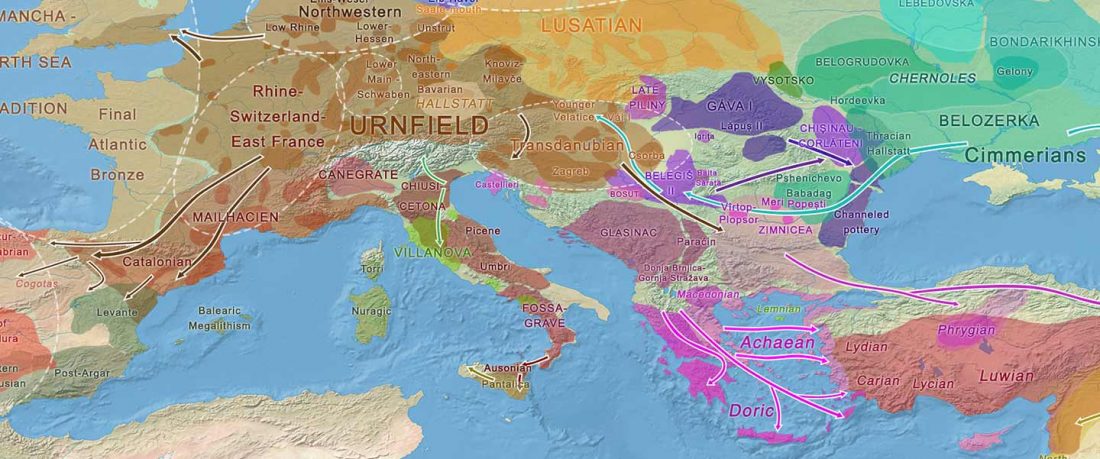
NOTE. I have updated prehistoric maps with Y-DNA and mtDNA data, and also the PCA of ancient Eurasian samples by period including the recently published samples, now with added sample names to find them easily by searching the PDFs.
Apennine homeland problem
The traditional question of Italic vs. Etruscan origins from a cultural-historical … Read the rest “R1b-L23-rich Bell Beaker-derived Italic peoples from the West vs. Etruscans from the East”