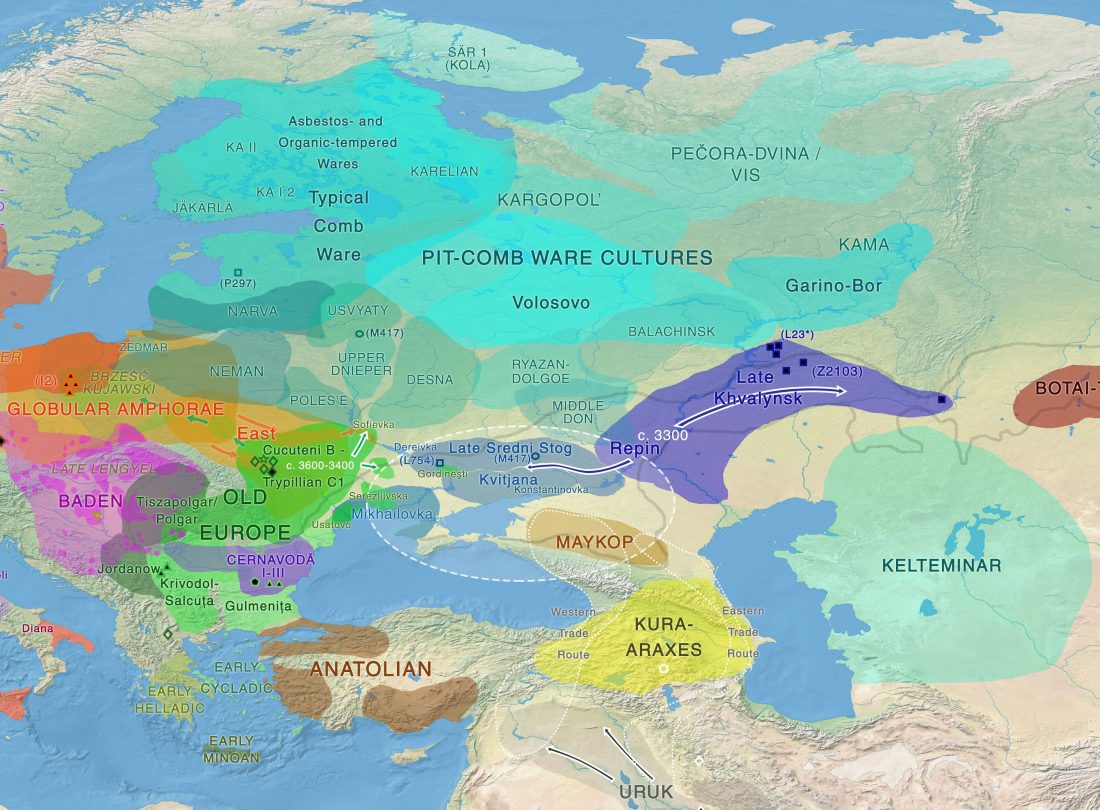
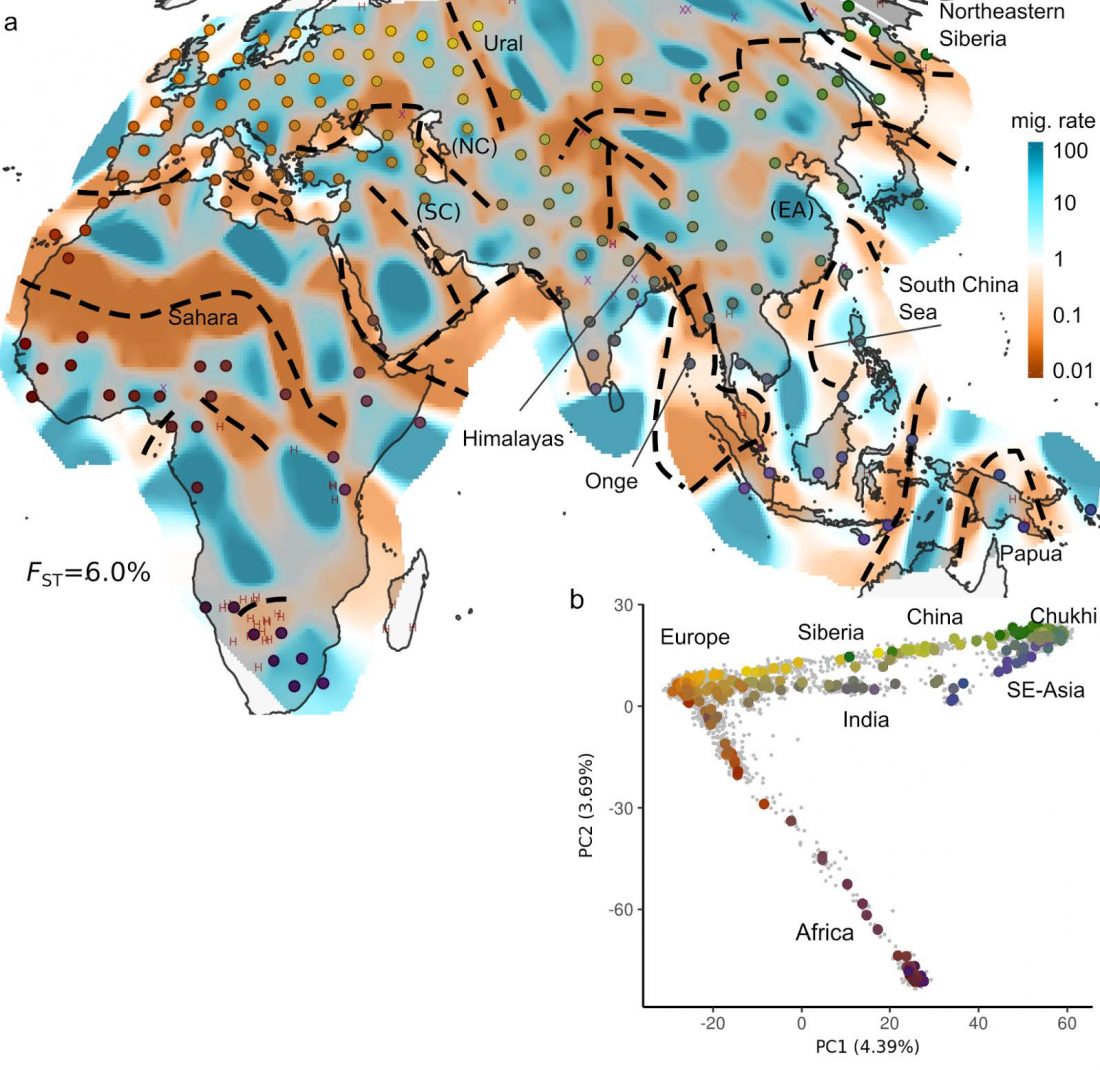
Open access Bayesian estimation of partial population continuity by using ancient DNA and spatially explicit simulations, by Silva et al., Evolutionary Applications (2018).
Abstract (emphasis mine):
… Read the rest “Bayesian estimation of partial population continuity by using ancient DNA and spatially explicit simulations”The retrieval of ancient DNA from osteological material provides direct evidence of human genetic diversity in the past. Ancient DNA samples are often used to investigate whether there was population continuity in the settlement history of an area. Methods based on the serial coalescent algorithm have been developed to test whether the population continuity hypothesis can be statistically rejected by analysing DNA samples from the same region but of different ages. Rejection of