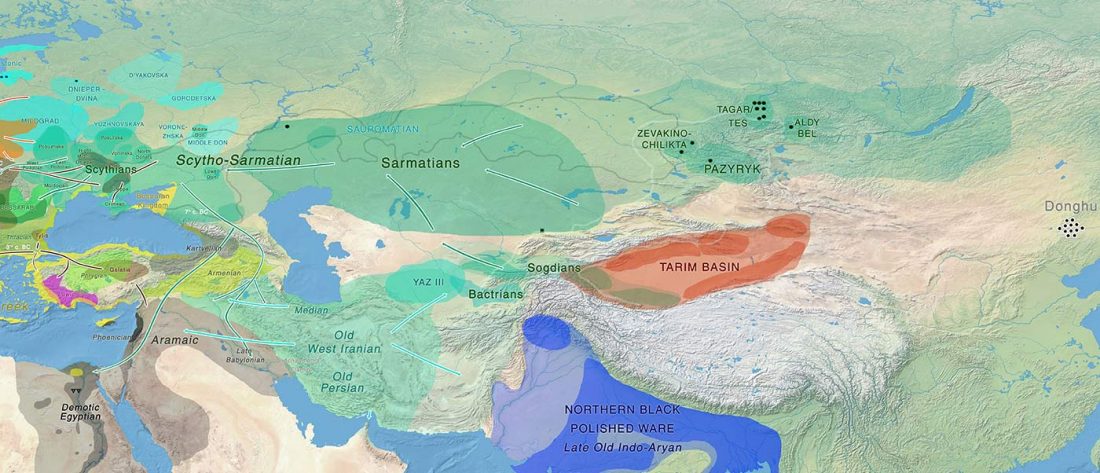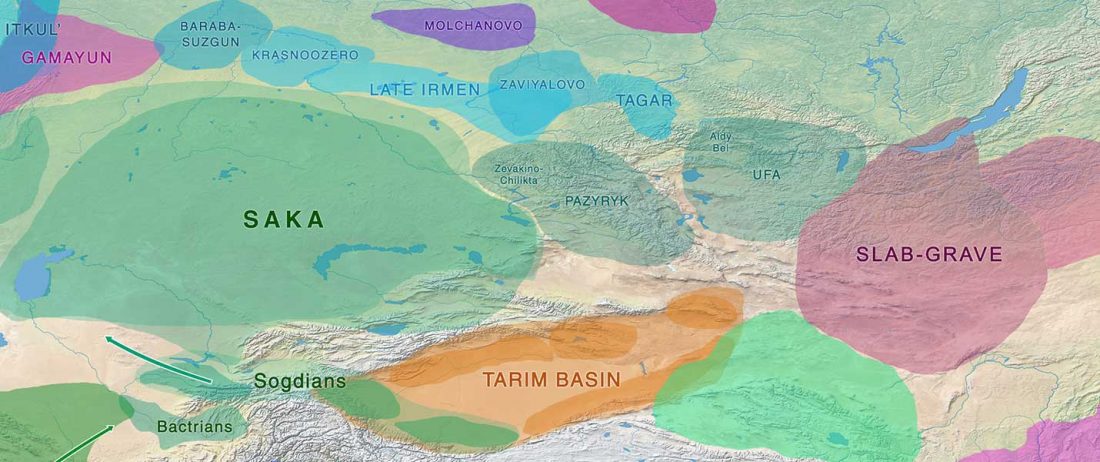
Recently, a paper described Eastern Scythian groups as “Uralic-Altaic” just because of the appearance of haplogroup N in two Pazyryk samples.
This simplistic identification is contested by the varied haplogroups found in early Altaic groups, by the early link of Cimmerians with the expansion of hg. N and Q, by the link of N1c-L392 in north-eastern Europe with Palaeo-Laplandic, and now (paradoxically) by the clear link between early Mongolic expansion and N1c-L392 subclades.
A new paper (behind paywall) offers insight into the prevalent presence of R1a-Z93 among eastern Scytho-Siberian groups (most likely including Samoyedic speakers in … Read the rest “Scytho-Siberians of Aldy-Bel and Sagly, of haplogroup R1a-Z93, Q1b-L54, and N”