Open access Genomic and Strontium Isotope Variation Reveal Immigration Patterns in a Viking Age Town, by Krzewińska et al., Current Biology (2018).
Interesting excerpts (emphasis mine, some references deleted for clarity):
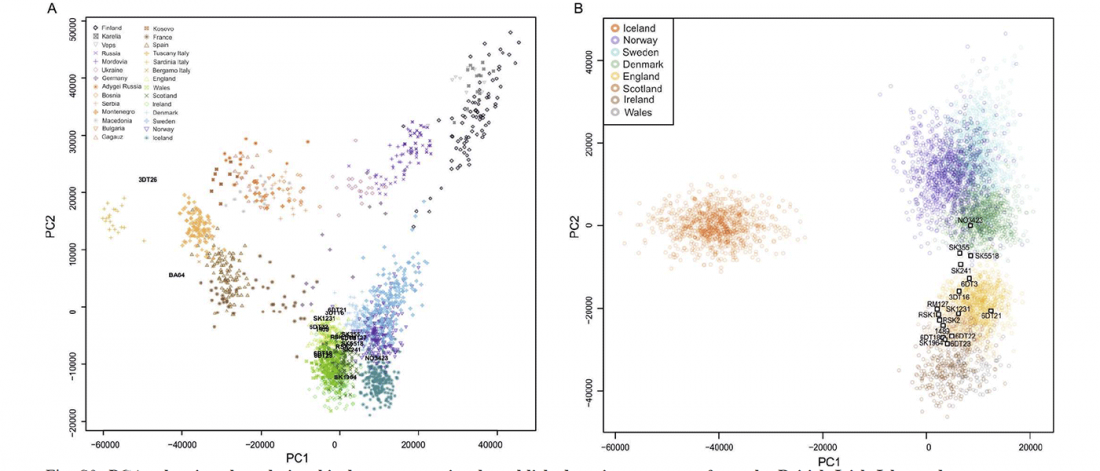
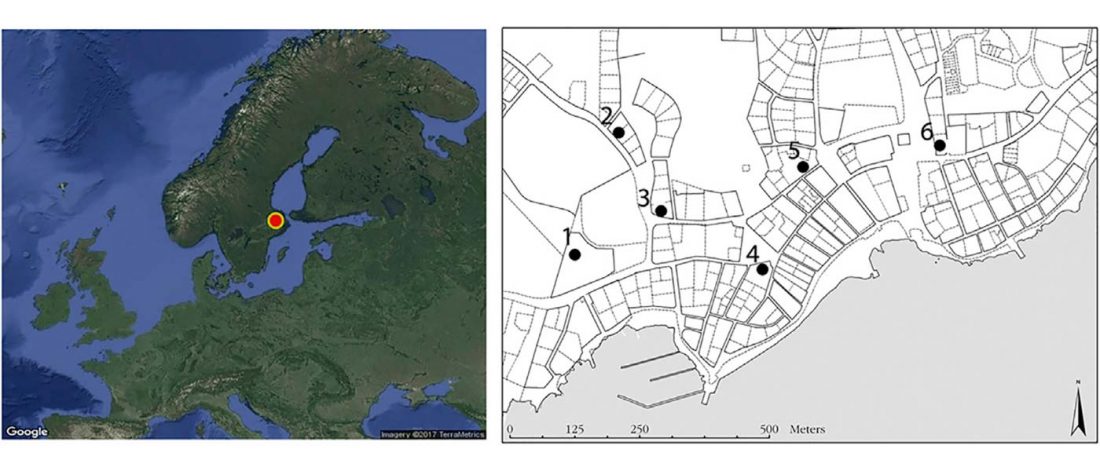
… Read the rest “Viking Age town shows higher genetic diversity than Neolithic and Bronze Age”The town of Sigtuna in eastern central Sweden was one of the pioneer urban hubs in the vast and complex communicative network of the Viking world. The town that is thought to have been royally founded was planned and organized as a formal administrative center and was an important focal point for the establishment of Christianity [19]. The material culture in Sigtuna indicates that the town had intense