This post is part of a draft on palaeolinguistics and the Proto-Uralic homeland. See below for the color code of protoforms.
7. Animal Husbandry
7.1. Milk
PFU (Saa., Fi., Md., Ma., P, Kh., Ms. Hu.) *waji ‘butter/grease’ (UEW Nº 1156).
NOTE. Aikio (2019: 48) contra Häkkinen (2009: 28-30) considers an original meaning ‘grease’ more likely than ‘butter’. Indeed, a more general meaning is always the most conservative approach. In fact, it was possibly borrowed from Smy. into PTurk. *bań ‘fat, grease; edible oil, butter; lubricant’ (Czentnár 2015: 9-17). What is striking about Aikio’s comments on this and the many doubts about other words related to Neolithic subsistence economy is that he himself (Aikio 2019: 46) doesn’t follow the safest alternative in many other, much more controversial cases, such as “Siberian pine”, “black-throated loon”, or “ski” (see above Foraging & Animals and Forests & Climate), so it seems that anthropological preconceptions occasionally prevail over his linguistic analyses. It is clear from many reconstructible words in this very text – whether they are natively Uralic or early borrowings – that early Uralic speakers had a (sub-)Neolithic subsistence economy. Furthermore, there is another word that covered the meaning ‘fat’ – PU (Fi., Md., Ma., P, Hu.) *kuje (UEW Nº 382) – which strengthens the complementary meaning of *waji.
PFP (Fi., Md., Ma.) *lüpsä-/*lupsa-/*lü(p)ś-tä- ‘to milk’ (UEW Nº 1396), suspected by Aikio (2015: 46) to belong to the Agricultural Substrate based on its “distribution, meaning and phonological irregularity”(?).
PCU (Ma., P) *liśtV- ‘to milk’ (UEW Nº 1385).
PFU? (Fi, Md., Hu.) *pejmä ‘milk’ ← PIE/Pre-PIIr. *peimn̥- ‘thick fluid; milk’ with a pattern similar to *tejniš (v.i.). Possibly the Pre-PMd. and Pre-PHu. verb ‘to milk’ from PIIr. *payas- ‘milk’ instead, cf. Av. paiias-, O.Ind. payas- (Holopainen 2019: 178-180 with references).
Pre-PFi *terneš ‘milk of a cow’ ← Pre-PIIr. *terun (from *ter- ‘soft’, hence ‘tender’) → PIIr. *taruna- ‘young of animals’, cf. O.Ind. táruṇa- ‘young, fresh’, Av. taᵘruna- ‘young of animals’.
PFP? (Fi., Md., Ma.) *utarV ‘udder’ (UEW Nº 1671) ← P(I)Ir. *ū́dʰr̥- from PIE *(h₁)ówHdʰ-r̥ ‘udder’. Divergent Uralic protoforms suggest parallel borrowings Fi.-Md. vs. Ma. (Holopainen 2019: 291-292 with references).
NOTE. Holopainen considers that a parallel loan in Mari would leave a possible PWU protoform *uδarV ← PGmc. **ūðara-/**ūðira- (since *δ would disappear in Mari), despite the rarity of Germanic loans reaching Mordvin. The actual forms proposed by Kroonen (2013: 129), which he references, are heteroclitic and amphikinetic r/n-stems PGmc. *eudra-/*ūdra-, though.
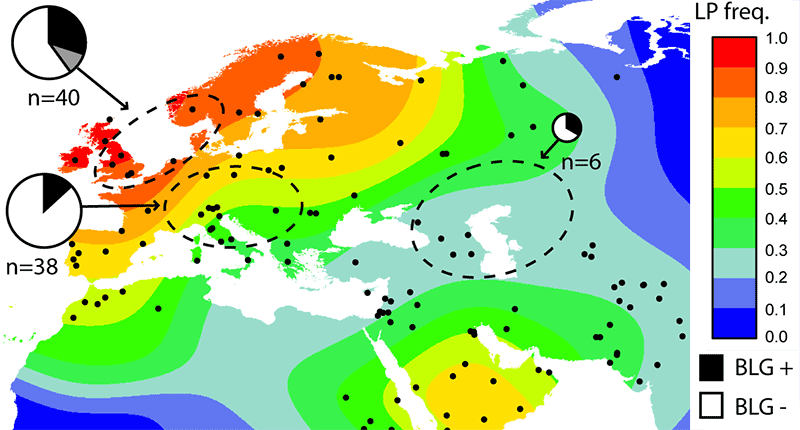
Milk products and their consumption in prehistory can only be inferred from indirect data (like wool, v.i.), such as lipid residues from ceramic vessels (e.g. Kučera et al. 2018), estimations based on weaning age of calves, etc. (e.g. Shukurov et al. 2015), or dental calculus (e.g. Warinner et al. 2014). While there is no detailed chronological map about specific cultures, it seems like milk product consumption spread with the Neolithic package, well before the known lactase persistence alleles (see recent papers on Europe and Africa, and a chronological map for ancient DNA). To the east, Afanasievo incomers ca. 3000-2500 BC represents the most likely arrival of dairy consumption in the Altai (Wilkin et al. 2020).
7.2. Ovicaprids
PFU (Fi., Md. P. → Ma.? Kh. ↔ Ms.) *uwčV/*owčV ‘sheep’ (UEW Nº 1086), difficult to reconstruct due to anomalies in sound correspondences; possibly a borrowing PKh. ↔ PMs. because of the irregular cognates PKh. *ā ~ PMs. *ā (Aikio 2020: 80-81 with references). Due to its irregularity, potentially an early Agricultural Substrate word with a widespread distribution, but in this case internal borrowings seem the most likely explanation.
NOTE. It has been proposed to be ultimately a cognate of PIE *owi- ‘sheep’, either from an unattested palatalized derivative in **-či, or from a common Indo-Uralic form, both alternatives unconvincing. Aikio (2015: 45) reconstructed Pre-PFi. *uwši for PFi. *ūhi, since the proposed development *č →*š →*h would still be irregular. However, South Finnic *utti, Pre-PMd. *o(w)ča, or Pre-PP *u(w)čV/*o(w)ča/*a(w)čV, apart from PKh. *āč and PMs. *āš, suggest an original *č as more likely.
Pre-PMd. *śaγa ‘goat’ ← PIIr. *sćāga-/*sćaga-, cf. O.Ind. cʰā́ga-, Oss sæγ (Holopainen 2019: 240-241 with references). Probably an Asian Agricultural Substrate word (Lubotsky 2001:313).
Pre-PP *warna/*we̮rna ‘wool’ ← P(I)Ir. *wárnā. Permic vocalism shows that the borrowing must be quite old (Holopainen 2019: 298-299 with references). From the same PIE word is the later borrowing PFi. *villa ‘wool’ ← PBal. *vìlna.
Later is the adoption of Pre-PP *mešV ‘ram’ (UEW Nº 1415) ← PIr. *mēš(a)- ‘ram, sheep’, from PIIr. *maiša-, cf. Av maēša- ‘ram’, maēšī- ‘sheep’, O.Ind mēṣa- ‘ram’, mēṣī́- ‘sheep’. Adopted from PP *mež into Mari (Holopainen 2019: 142 with references).
Sheep-Goat and Wool in Archaeology
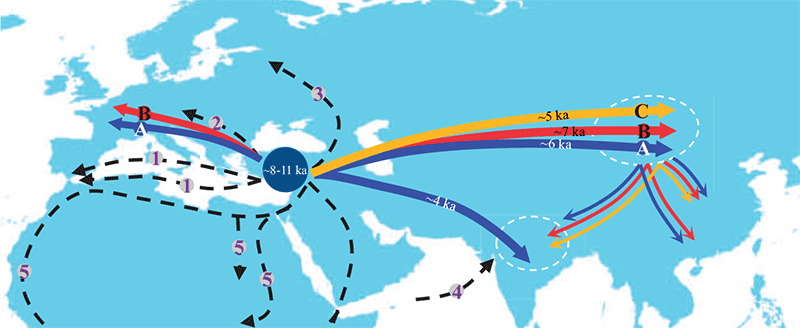
Sheep and goat husbandry appear to stem from Iran, and the technique spread quickly to the Caucasus and to Europe with the Neolithic expansion (Alberto 2018). Nevertheless, there are still doubts about the specific chronology and lineages involved in each territory. Particularly relevant for the PIE and PU question is their spread to the Altai, which seems to have happened first with the expansion of Pre-Tocharian-speaking Afanasievo at the end of the 4th millennium BC (Hermes et al. 2020). It can be assumed that ovicaprids and cattle spread to North-Eastern Europe and beyond the Urals first with the Corded Ware culture and its later offshoots, with some Y-chromosomes extant today potentially associated with that expansion (Deng et al. 2020). A parallel evolution can be inferred for goats (Zheng et al. 2020).
Wool economy boomed in continental Europe mostly during the Bronze Age (Sabatini 2019). In archaeology, wool sheep finds might be indirectly assessed through the shift towards sheep husbandry (and related proportionally to goats); slaughter patterns (more males are kept alive until adulthood); appearance of smaller and lighter spindle whorls to spin short wool fibres; identification of tools for wool dyeing; iconography of garments (although plant fibre garment is not easily distinguishable from animal fibre garment); direct wool textiles; Bronze Age texts, etc. (Strand, Nosch 2019)
The earliest known Eurasian textile fragments date back to the 7th-6th millennium BC and are made from vegetable fibres, with domestic flax cultivated in the Mediterranean, SW Asia and Caucasia since before 6600 BC. Woollen textiles are a later invention, with the first likely evidence found in ancient written texts from Mesopotamia during the 4th millennium BC (Sabatini, Bergerbrant 2019). Nevertheless, following indirect data, Anthony (2007) considers sheep wool to have been present by the Late Neolithic TRB in upland southern Poland by ca. 3600 BC, based on the number of adult caprids. In the Northern Caucasus, it is likely to have appeared earlier (ca. 4000 BC). Only the neighbouring steppes and South Asia might have also been prolific in sheep wool herds during the Final Eneolithic / Early Bronze Age.
Outside of these areas, the Usatovo economy among Late Trypillians (as inferred from Usatovo and Mayaki materials) seems the most prolific in ovicaprid economy, and potential sheep wool butchering in particular, probably due to their location in the steppes and through intense contacts with the Yamnaya. There are also findings of spindle whorls and horses (also shown in horse images), and its territory in the middle of chariot-building Yamnaya/early Catacomb herders suggests that it could have been a vector of the earliest Indo-European and Pre-Indo-Iranian loans into the forest-steppes and forests of Eastern Europe (see below Textiles in Technology)
7.3. Cattle
PFP (Md., Ma., P) *skal ‘cow’ (UEW Nº 1669), with some Ma. and P forms reflecting a prothetic vowel – hence the traditional reconstruction *uskalV – maybe due to contamination with *uška (v.i.). Possibly a borrowing due to the irregular sound correspondences (Aikio 2014: 128).
PFP (Fi., Ma.) *tejniš ‘pregnant (of animals)’ ← Pre-PIIr. or (Pre-)PBSl. from PIE *dʰe-in-, found in PBal. *deini (cf. Lith. diení ‘with young’, Latv. atdiene ‘a cow that calves already in its second year, calf that has not yet produced milk’), and PIIr. *dʰainúš (cf. O.Ind. dʰenú- ‘cow, milking cow’, Av. daẽnu ‘female, suckling’). Semantically more attractive from (Pre-)PBSl. into Finno-Permic, but could be from PIE or Pre-PIIr. just as well, or even separate borrowings in Pre-PFi and Pre-PMa (Holopainen 2019: 276-277 with references).
NOTE. Compare also Pre-PHu. *täjVnV ‘cow’ from PIr. *daynu- ‘cow’ (Holopainen 2019: 287-288 with references); Pre-PHu. *täji/*täjä ‘milk’ ← Alanic, cf. Oss dæjyn, dæyun ‘(breast) sucking’ (Holopainen 2019: 288 with references).
PFP (Pre-PP → PMa. – Ms.?) *uška ‘ox, bull’ (UEW Nº 1670) ← PIIr. *ukšan-, much less likely from PIr. *uxšan-, which would have rendered Pre-PP **uša or the like (Holopainen 2019: 290-291). However, the PIr. form is more likely behind Proto-Kartvelian (also metathesized) *usx- ‘sacrificial bull’ → Geo. usxi, Svan usxway ‘sacrificial bull’, as strongly suggested by cultural evidence of bull-slaughtering rituals in the Gathas, Yasna 29.1 & Yasna 29.6 (Klimov 1994: 64-58; Fenwick 2017: 4; Bjørn 2017: 113).
PFP (Fi., P) *ćońi ‘young male animal (foal, calf, bull)’ (UEW Nº 1205). Irregular vocalism in Permic (should point to PP *o̭) suggests a potential borrowing.
PWU (Fi., Md.) *warsa/*we̮rsa ‘calf’ ← PIr. *wŕ̥šan- as a “Pre-Alanic” form, cf. Alanic *wәrsa- behind Oss wyrs, urs (Holopainen 2019: 299-300 with references).
PWU (Fi., Md.) *wasa/*we̮sa ‘foal’ (UEW Nº 1691) ← PIr. *wasá- (← PIIr. *watsá-), cf. Oss wæs ‘calf’ (Holopainen 2019: 300-301 with references).
PFP (Saa., Ma., P) *počaw ‘reindeer’ ← PIr. *pacu- ‘cattle; livestock’. Most likely a wide Eurasian Wanderwort; cf. parallel loans POUg *pǟčäγ ← PIr. *pacuka, and possibly also behind Fi and Ka poro, and many other unrelated languages in Northern Eurasia (Holopainen 2019: 184-185 with references). The source language(s) and originally designated animal(s) in each case are unknown.
NOTE. While conclusive evidence about the lack of reindeer in or close to the Uralic homeland is impossible – after all, vocabulary for the more widely distributed deer or moose seem to be interchangeable for reindeer throughout language history – it is particularly telling that vocabulary for reindeer and reindeer herding would need of an early Iranic (or later, indirect Siberian) loan, which seems to support the proposed late emergence of the technique, after 1500 BC or later during the Early Iron Age (see e.g. Pelletier et al. 2020). Compare also West Uralic *wasa/*we̮sa ‘calf’ → ‘elk calf’ above.
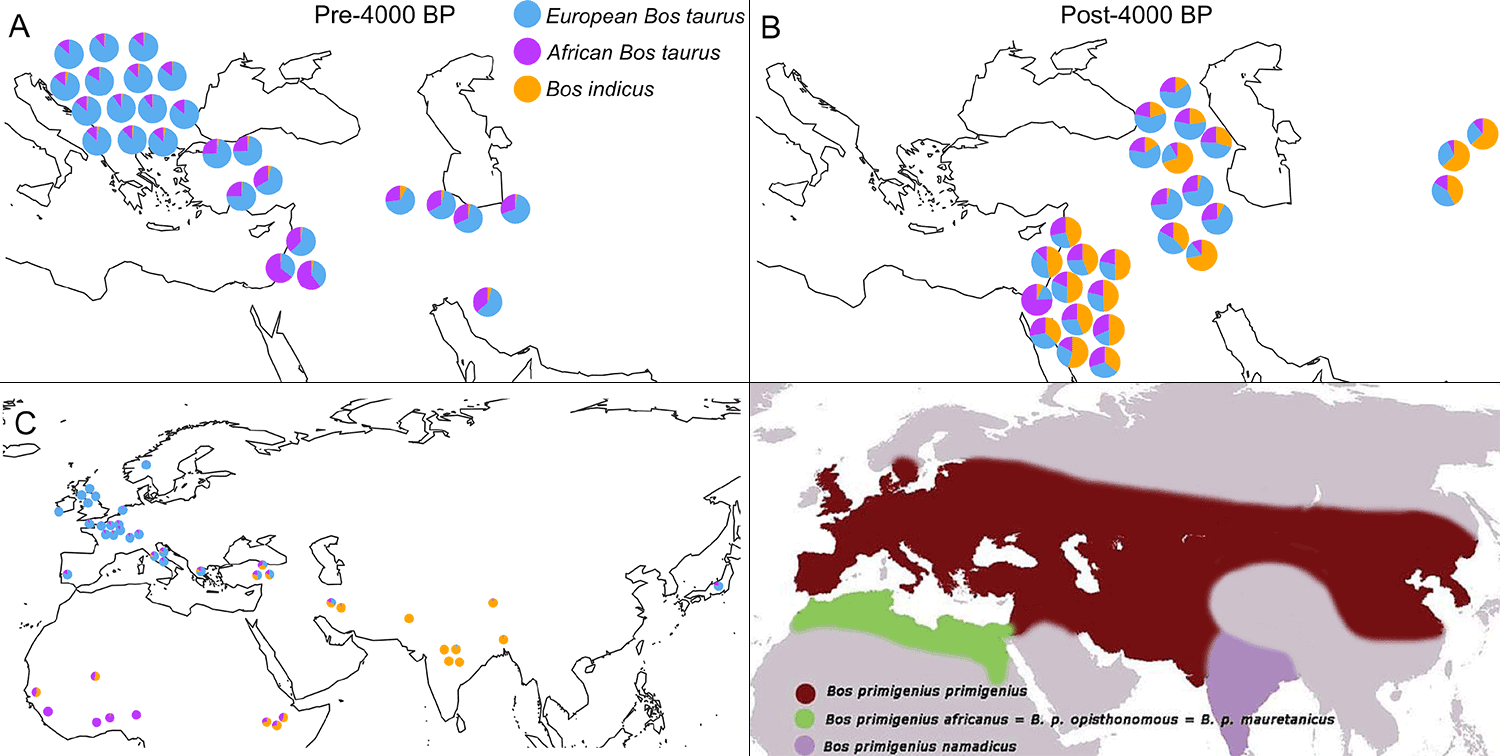
Cattle were domesticated from wild cattle. It is generally thought that cattle was domesticated independently twice in two different geographical locations: in Anatolia and the Indus Valley (Loftus et al., 1994). This led to the two major groupings of domestic cattle (Scanes 2018), with B. taurus taurus (or European type cattle) being present in the Early Prepottery Neolithic (ca. 8800-8300 BC) in the Fertile Crescent, in Southeastern Europe by ca. 6700 BC, Southern Italy by ca. 6500 BC, Central Europe by ca. 6000 BC, and in Northern Europe with cheese being made as early as the 6th millennium BC (Salque et al. 2013).
Cattle herding started early in the steppes (Morgunova et al. 2019), and probably spread to the Altai with Yamnaya-Afanasievo, and to North-Eastern Europe and the southern Urals probably first during the Early Bronze Age with the Corded Ware culture. B. taurus indicus and their hybrids are known to show markedly enhanced production in arid and thermally stressed environments compared to B. taurus taurus, which explains their later expansion in the Middle East (Pereira Verdugo et al. 2020).
{kind=link}
7.4. Horse
PWU (Saa., Md.) *ältV ‘mare?’ (UEW Nº 1194). A regular Uralic cognate (Aikio 2020: 36-37 with references), it was borrowed from Pre-PSaa. *āltō → Pre-PGmc. altō-, cf. Proto-Nordic *eldō- in O.N. jalda ‘mare’ (Hofstra 1997: 127–129), which hints at its ancestral meaning in Saami, ‘mare’ (today ‘reindeer cow’), in common with PMd. *äľďǝ ‘mare’.
NOTE. Due to the phonological evolution of Germanic, it must be a very old borrowing from Pre-PSaa. or both PWU and Pre-PGmc. from a third, unknown language (Aikio 2020: 36-37). According to Aikio (2002: 23), the presence of regular counterparts in Samoyedic and the lack of a good Indo-European etymology points to this direction of borrowing. Other early borrowings supporting this are PWU *muδ́a and PFi. *mato, both borrowed into Pre-Grimm Germanic (see above). Aikio’s proposal of a (theoretically conceivable) borrowing from a third language concerning the Uralic and Germanic cognates – justifying it on the scarcity of Uralic loanwords in Germanic – cannot hold on linguistic grounds alone, since the PSaa. ~ PMd. cognates are regular, and so are the PSaa. ~ Pre-PGmc. cognates, hence it risks rejecting multiple solid evidences based solely on (anthropologically-based?) circular reasoning.
PWU *lešmä ‘large domestic animal’ (UEW: 689), cf. Fi. lehmä ‘cow’, Md.E. ľišme ‘horse’, probably from a substrate language (Häkkinen 2009: 38; Aikio 2015:45; Zhivlov 2015; Holopainen 2019: 265 with references). Based on PFP *skal ‘cow’ (v.s.), possibly more likely to have referred originally to ‘horse’.
PUg. (Ms., Kh. → Smy., Hu.) *luwV (*luxe) ‘horse’ (UEW Nº 1794). Possibly related to PToch. *lŭwā-(ke) ‘animal’ (←? PIE *luhₓeh₂) or PSlav. *lȏvъ ‘hunt, hunting; game animal?’ (←? PIE *louhₓo-), hence to the assumed parent IE protoform – maybe also behind Gk. léōn ‘lion (←? predator)’ (Adams 2013: 606-607). No closer – chronologically and dialectally more attractive – (Pre-)Indo-Iranian cognates can be reconstructed, even though semantically it would still be easier to accept an earlier borrowing (“horse as game”?; maybe “horse as domesticate”? v.i.) rather than from specialized horse-riding and chariot-driving (Pre-)Proto-Indo-Iranians. In any case, due to the early and short period of the Ugric proto-language or Sprachbund, a quite early Uralic (loan?)word.
NOTE. Generally assumed to be a loanword because of its meaning and distribution rather than on purely linguistic grounds:
- It could fit the (loosely described) intervocalic laryngeal correspondence (Pre-)PIE *H ~ (Pre-)PU *x (see below Earliest External Contacts), Pre-PU **luxe̮ ~ PIE *leuhₓ- ‘cut off, separate’, the most likely root behind PIA *luhₓp- ‘hide, skin, flay’ (Adams 2013: 606-607), which would connect the term with domesticates. On the other hand, a connection of IE *lāp with Semitic *álap- ‘bull, cattle’ has been proposed (cf. Bjørn 2017: 91).
- Based on the alternating *-m-/*-w- of the Agricultural Substrate language (Zhivlov 2015) and the generalized lack of its characteristic *-š-loanwords in Ugric, potentially a cognate of *lešmä in Ugric; however, neither a clear protoform nor similar examples exist to compare to.
- Keresztes’ (1998:416) proposal of a Turkic origin does not hold.
- Parpola’s (2012: 296) proposal of an origin in the Botai cannot be formally tested – lacking any other believable example – and is chronologically unattractive, since their demise predates Proto-Indo-Iranian-speaking Sintashta-Petrovka (assumed to have been in contact with early Proto-Ugric dialects) by more than 1,000 years.
- The proposal by Zaicz (2006: ló[1055]) of a borrowing from a Caucasian language (viz. Avar thah ‘stallion’) is left unexplained.
Later loans are e.g. Pre-PP *ačwa/*e̮čwa ‘stallion’ ← PIr. *ácwa- (← PIIr. *áćwas) ‘horse’, cf. Av. aspa- (Holopainen 2019: 54-55 with references); and Pre-PP *parśa/*barśa ‘horse’s mane’ ← Alanian *barci- or PIr. *barša- (Holopainen 2019: 71-72 with references).
NOTE. On PFi. *hepoi ‘horse’ ←? Pre-PFi. **šepä? ~? PIE h₁ekwos (Katz 2003), there are no uncontroversial examples of PU *š ← PIE *H, and it is unclear how the cluster PIE *-kw- or (N)WIE *-kʷ- would have been adopted. It cannot go back to Pre-PGmc. *ekʰʷoz and even less PGmc *ehʷaz for loss of initial laryngeal well before that date (or, at least, generalized lack of effect of *H in Uralic borrowings). The proposal of a relationship with a source akin to ON hoppa ‘jump’, from PGmc *huppōną is phonologically and semantically uncompelling.
Tracing the spread of the domesticated horse is important to understand the Proto-Indo-European expansions. However, while there can be little doubt that the Proto-Uralic community knew the horse – based on its prehistorical (wild and domestic) range and the combination of all other cultural and economic vocabulary involved, see below – it is unclear whether early speakers of Uralic dialects handled or even rode domesticates.
7.5. Pigs and boars
PWU (Saa. ← Fi., Md.) *tika/*toka ‘swine’ (UEW Nº 1644), suspected by Zhivlov (2015) and Aikio (2015:46) to belong to the Agricultural Substrate based on its “distribution, meaning and phonological irregularity”(?).
PWU (Fi., Md.) *woraći/*woraća ‘pig, boar’ (UEW Nº 1458) ~ PIIr. *warāʒ́ʰá- ‘boar, swine’ (Parpola 2005: 42-45). Usually assumed to be a borrowing in Indo-Iranian from the Asian Agricultural Substrate, the regularity between both West Uralic branches hints to an early Uralic borrowing (Holopainen 2019: 313-314 with references).
PU (Fi., Md., P) *porćas/*porśas ‘pig(let)’ (UEW Nº 1458) ← Pre-PIIr. porćos / PIIr. *parća-s (cf. Av pərəsō, Khotanese pāsa-) or PBSl. *porśo-s. Mordvin belongs probably to the same protoform as Pre-PFi., but the irregular Permic vocalism might point to an Early Iranian loan, or alternatively an influence of *r (Holopainen 2019: 190-193 with references). A PIIr. borrowing among already split-up Uralic dialects seems a priori more likely than parallel PBSl. + PIr. borrowings.
? Pre-PFi. *kärsä ‘snout’ ← P(I)Ir. *karṣū́- ‘furrow’. The borrowing could be related to the boar’s use of its snout to root around in the ground (Holopainen 2019: 129-130 with references).
PU (Saa., Fi., Smy.) *tora- ‘fight’ ← PIE/Pre-PIIr. *dʰor- / PIIr. *dʰār- (Holopainen 2019: 282-285). One of the few (Pre-)PIIr. loanwords with a wide distribution stretching to Samoyed. It possibly belongs here due to the later extension into PFi. *torakko, *torikko ‘tusk of wild boar’ (see below).
Celtic substrate
The work by Hyllested (2012) shows conceivable correspondences between Pre-Proto-Celtic and Pre-Proto-Balto-Finnic, such as:
- PCel. *mokku- ‘swine’ ~ PFi. *emakko, *emokke ‘sow’.
- PCel. *sukko- ‘sow’ ~ PFi. *cuka ‘swine’ (→ Ltv. cūka ‘pig’).
- PCel. *turko- ‘wild boar’ ~ PFi. *torakko, *torikko ‘tusk of wild boar’, analyzable as derivative from PU (Saa., Fi., Smy.) *tora ‘fight’, in turn from PIE *dʰor- (→ PIIr. *dʰār-).
NOTE. Matasović (2008: 359) mentions a potential analogical influence of PCel. *mokku-, but also that a borrowing from an unkonwn source remains a possibility. The WGmc. forms in -k- can be explained by assuming levelling from the accusative case, and in Nordic it rose through Holtzmann’s law (Kroonen 2013: 490). The existence of geminated derivatives supposedly from a reconstructible PIE *suhₓ-ko- in Indo-Iranian makes Hyllested’s interpretation a priori less attractive. However, taking the common NWIE stage and laryngeal loss into account, the evolution of the form in Celtic is fully unexplained.
Hyllested proposes that these shared terms and their significance for Celtic and Germanic mythology may emanate from a common, non-IE source. This case would be similar to the origin of ‘seal’ in both Germanic and Balto-Finnic, with an initial adoption by the latter (see above). As indirect support for this, he mentions:
- PCel. *brokko- ‘badger’ an animal usually conflated with pigs, is also found in W.Gmc. *brakka- ‘(scent) hound, dog used for hunting’, hence both possibly from the language of the geminates (Schrijver 2001).
- PIE *porko- ‘pig’, a NWIE term shared with N. Iranian, might be also proposed as a substrate word (including Uralic forms) due to the initial *p and the need to reconstruct irregular variants such as *porgo- and *bʰorku(o)- or *bʰorgʰu(o)- for Germanic and Slavic forms.
Holopainen (2019, passim) uses, again, anthropological preconceptions and circular reasoning to reject these proposals: “it is very unlikely to postulate a contact situation where [*tora] was borrowed into Proto-Celtic (!)”; “[i]t seems very improbable that there were any kinds of contacts between Celtic and Finnic at any time”; “there is no (paleolinguistic) evidence that speakers of early varieties of Uralic would have practised pig herding”.
However, the whole set of early Uralic pig/boar related vocabulary relative to the Proto-Indo-European one suggests that pig herding was, in fact, as important for the subsistence economy of early Uralic peoples as it was for early Indo-Europeans (who also adopted the words from neighbouring peoples), and that West Uralic was in immediate contact with Pre-PGmc. and Pre-PBSl, and both PU and PIE probably (indirectly?) with similar substratal Neolithic language(s). In fact, there are PWU loanwords in Celtic for ‘pine’, ‘ash-tree’, ‘mulberry’ (see above), or PCel. *wē(t)tā ‘stream, swamp’ ← PWU *weti (de Smit 2019: 8) – the latter maybe also through the language of the geminates (Schrijver 2001, 2005) – which support such early (indirect?) contacts.
Pig domestication
The origin and spread of pig domestication in Eurasia appears to be roughly shared with that of ovicaprids and cattle. However, its start and evolution seem to be more complex, with an interaction with humans and admixture with their wild relatives comparable to that of early domestic horses with wild ones (Price et al. 2020). In archaeology, theories of its presence and relevance are therefore slightly more difficult than for other species (Price & Hongo 2019). Around the steppes, the picture is not easier, either: many sites from steppe regions like Azov, Northern Caucasus, West Caspian/Kalmykh, North Caspian/Ural steppes are essentially devoid of pigs, whereas sites from forest-steppe ecotones of Trypillia, Sredni Stog, or Abashevo show a non-negligible proportion of pig domesticates (and/or boars). Indirect data about domesticated pigs might also be inferred from the relevance of agriculture in the subsistence economy.
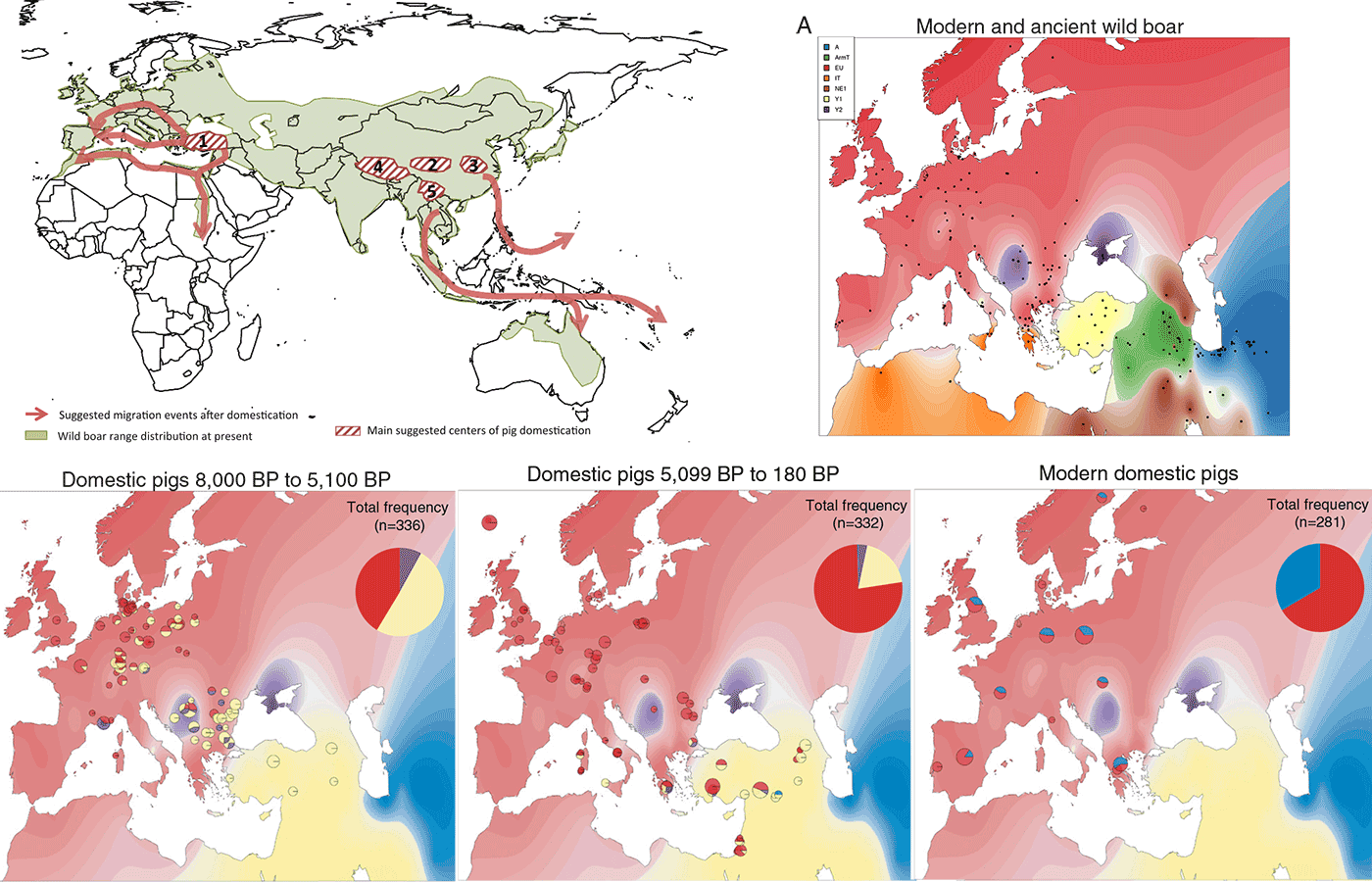
Top Right: The black dots on the map represent locations of 696 modern and ancient wild boar that possess East Asian (blue), Near Eastern (including haplogroups mt-Y1 and mt-ArmT; yellow), European (including haplogroups mt-Italian, mt-A, mt-C, and mt-Y2; red), and Y2 (purple) haplogroups. The haplotype assignments were used to interpolate the underlying color distribution which demonstrates the biogeographical boundaries of these 3 general haplotypes. Image modified from Franz et al. (2019) Supp. Materials.
Bottom: (Left) Large pie chart in the upper right corner of the map represents overall frequencies of these haplogroups in domestic pigs. Small pie charts on the map show the frequencies at various archeological sites/locations between 8,000 y BP and 5,100 y BP (B), between 5,099 and 180 y BP [before the Industrial Revolution and the introduction of Asian pigs in Europe (Center), and in modern pigs (Right)]. Images modified from Franz et al. (2019).
7.6. General
PFU (Fi., Md., Ma., Hu.) *wetä ‘lead, guide’ (UEW Nº 1141) ← PIE/NWIE *wedʰ-e- ‘lead, guide’, found with an e-stem in NWIE (cf. O.Ir. fedid, Goth. ga-widan, Lith. vedù, OCS vedǫ), but with an o-stem in Hittite and Indo-Iranian (LIV²: 659).
PFP (Fi., P) *tarna/*te̮rna ‘grass’ (UEW Nº 1632) ← PIIr. *tŕ̥na- ‘grass, blade of grass, herb’, cf. PFi. *tarna, PP *turi̮n. It could also be from a Pre-PIIr. or earlier *tŕ̥no-, or even a later, PIr. borrowing, but possibly “connected to the influence of a pastoral culture of the Indo-Iranians” (Holopainen 2019: 273-274 with references), in light of the parallel borrowing of PIIr. *ándʰas in a more evident mythological context.
NOTE. To support this borrowing as related to steppe grasslands, cf. PFU (Saa., Fi., Kh., Ms.) *ńurmi ‘meadow / grassy bog’ (UEW Nº 649) and PFU (Fi., Kh., Ms.) *ara(w) ‘old river bed / oxbow lake (?)’ (UEW Nº 27) with derivative *ura- ‘groove’. According to Aikio (2020:27-28), the meaning ‘flood meadow, boggy grassland next to a river’ must be more archaic than ‘dry grassland’ for both terms.
PFU? (Saa., Kh.) *ćuri ‘herd’, reconstructed from PSaa. *ćorkē ‘herd of reindeer’ and PKh. *sōr ‘small herd of wild reindeer or moose’ (Aikio 2020: 143-144 with references). If this etymon is reconstructible for the parent language, then a further proof that Proto-Uralic speakers practised herding and had a word for it that was likely replaced through intense contacts with Proto-Indo-Iranians, i.e. well before the emergence of reindeer herding.
PFP (Saa., Md., Ma., P) *ert(t)ä ‘side, rib’ (UEW Nº 1227) ← Pre-PIIr. *érdʰo- / PIIr. *árdʰa- (cf. Skt. árdʰa- ‘side, part, half, place’, Av. arǝδa- ‘side, half’), or PIIr. *r̥dʰakás ‘side’; cf. O.Ind. adverb r̥dʰak ‘separately’ (Holopainen 2019:81-83 with references; Aikio 2020: 47-48).
PFP? (Saa., P?) *kert(t)ä- ‘to tie’ ← Pre-PIIr. *k(e)rt- / PIE kʷ(e)rt- (cf. PGmc. *hurþi-). PP *kerti̮- ‘to tie’ also potentially derivable from PU *käri- ‘wrap, tie’ (Holopainen 2019: 119-120 with references).
PFP (Ma., P) *wirta- ‘to raise, to hold’ (UEW Nº 1716, cf. PMa *wŭrδe-, PP *vordi̮-) ← PIIr. *wardʰ- or Early PIr. *ward-, PIr. várdʰati, from PIE *werdʰ- ‘to become big, strong’. From the set of loanwords related to cattle and horse breeding (Holopainen 2019: 311-312 with references).
Pre-Saa. *kata-/*kato-/*ke̮ta-/*ke̮to- ‘to graze’ ← PIIr. *k(ʰ)ād-, potentially also earlier PIE *kād- (unattested in Germanic or Balto-Slavic) or PIr. *xād-, but most likely Indo-Iranian (Holopainen 2019: 116 with references).
PUg.? (Ms., Hu.) *oćtVrV ‘whip’ (UEW Nº 658) ← PIIr. *aćtrā- ~ *aƷ́trā- ‘whip’, cf. O.Ind. aṣtrā-, Av. aštrā-. Different PMs. *āćtər and OHu. Ystwr support early parallel borrowings after the split-up of Proto-Ugric (Holopainen 2019: 156-157 with references).
PFU (Saa., Fi., Kh.) *šuwli ‘lip, mouth’ (UEW Nº 1012; Aikio 2015:44). Based on the reconstruction of an *š, possibly an Agricultural Substrate word, but with a widespread distribution in Uralic. It might have been originally related to animals.
PU? (Fi., Md., P, Ms., Smy.) *kajšo ‘sickness’, *kajši- ‘be sick’ (Aikio 2014: 3–5; 2015: 44; 2020/forthcoming). Based on its irregular reconstruction and the presence of an *š, possibly an Agricultural Substrate word, but with a widespread distribution in Uralic.
NOTE. It is particularly attractive to speculatively conflate this word and the recent presence of the plague in the Neolithic world, probably one of the reasons for the expansion of populations of “Steppe” ancestry. Of course, the word ‘sickness’ need not have referred to ‘the plague’. It is a mere speculation of an adoption of a foreign word based on indirect data: (a) the potential substratal nature of this widespread term, (b) its spread roughly around the Proto-Uralic chronological framework, and (c) the existence of many other “regular” P(F)U words for ‘ill(ness)’. On the other hand, it could speculatively be associated to an originally onomatopoetic formation of ‘sneeze’, which would make sense for those widespread words in *š (like *pušV- ‘blow’ UEW Nº 827). Such an use of this rare phoneme would be similar to the existence of PIE onomatopoetic laughing noun and verb in *kʰa- (*xa?) but few if any other PIE words with this phoneme.
References
See full Bibliography.
Color code for protoforms
- certain | uncertain
- PU/PFU | PFP/PWU/PFS/PFV/PUg.| Dialects
- PIE (PIU?) | PIE? (PIU??)
- PIE/Pre-PIIr. (PIU??) | Pre-PIIr. | PIIr. | PIr.
- NWIE | Pre-PGmc./(Pre-)PBSl. | PGmc./PBal./(Pre-)PSlav.
- Substrate | Substrate?
- Agricultural: PU | PU? | PIE | PIIr.
- Geminates | Eurasian (Wanderwort?)
- Palaeo-European | Palaeo-Laplandic/Arctic
Return to the top section: ↑ 7. Animal Husbandry.
Featured image: Erdal-Bilderreihen 5/125 Eisenzeit – Germanische Gehöft, by Gerhard Beuthner (1930?).