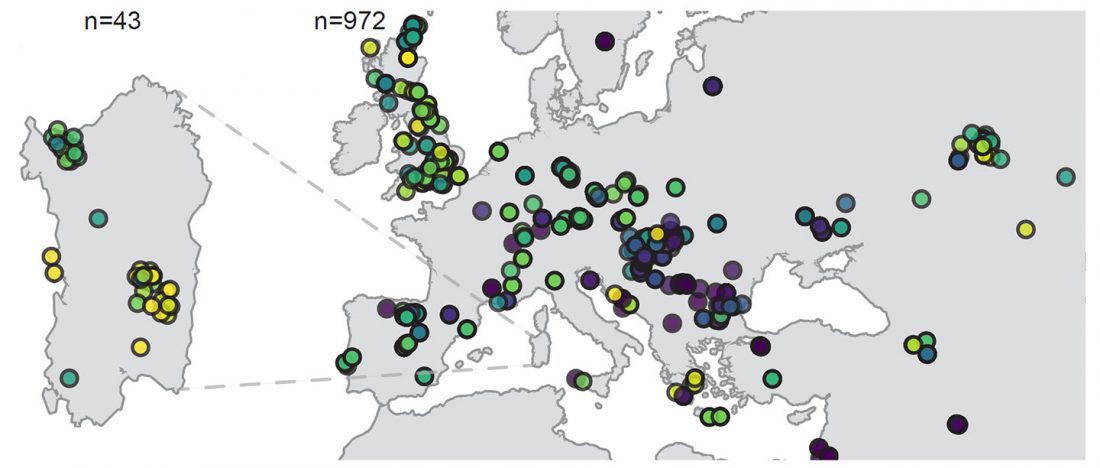
Recent papers on France and neighbouring regions, Ancient genome-wide DNA from France highlights the complexity of interactions between Mesolithic hunter-gatherers and Neolithic farmers, by Rivollat et al. Science Advances (2020) 6(22), and Ancient genomes from present-day France unveil 7,000 years of its demographic history, by Brunel et al. PNAS (2020).
Interesting excerpts (emphasis mine):
I. Survival of HG ancestry in Central Europe
From Rivollat et al. (2020):
Here, we present newly typed genome-wide data from 101 individuals from 12 sites from modern-day France and Germany (3 Late Mesolithic and 98 Neolithic, 7000–3000 cal BCE (…)
… Read the rest “Survival of hunter-gatherer ancestry in West-Central European Neolithic”We explored