Dynamics of paleoenvironments in the Cis-Ural steppes during the mid- to late Holocene, by Khokhlova, Morgunova, Khokhlov, and Golyeva, Quaternary Research (2018), 1–15.
Interesting excerpts:
About the studied site
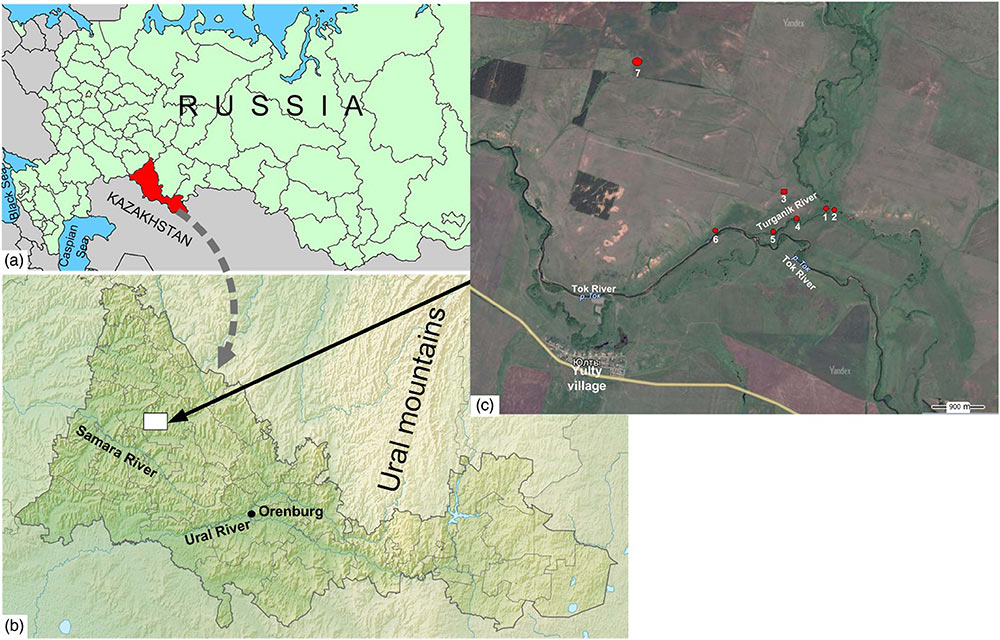
The Turganik settlement in the Orenburg Region constitutes part of the so-called Ivanovo microregion of cultural heritage monuments, along with the Mesolithic Starotokskaya site; an Ivanovskoye multi-layered settlement (Neolithic, Eneolithic [or Chalcolithic], Late Bronze Age); Toksky I and Toksky II settlements attributed to the Late Bronze Age (the Timber-Grave archaeological culture); an Ivanovsky ground burial dated to the Eneolithic; and the Ivanovsky kurgan cemetery of the Early Iron Age (Fig. 1).
The ancient settlements are located at the Turganik River mouth, where the river joins the Tok River (the Samara River drainage basin). The Turganik River enters an old channel of the Tok which continues to flow due to that fact. Both valleys are wide and dissected by multiple river channels. The floodplain landscapes are mostly wet meadows with rich herb and grass vegetation, pastures, and hay fields. On both sides of the Turganik River, and farther along the right side of the Tok valley there are flat-topped elevations, with occasional forests (Chibilev, 1996). The Turganik settlement was positioned on a slightly elevated surface at the confluence of the Turganik and Tok rivers, on the right side of the valley. The settlement was inhabited in the Eneolithic and the Late Bronze Age, the fifth to fourth millennia BC.
Results and discussion
Pollen assemblages of the Atlantic optimum ~ 5500 yr BP indicate some increase in moisture supply and related afforestation of the floodplain (Lavrushin and Spiridonova, 1995). As follows from our data, the site was abandoned at that time and the no-longer-functioning cultural layer VI was gradually buried under deposits of frequent floods. According to the 14C ages obtained on archeological materials, the age of layer VI (or the second stage of the Eneolithic epoch on the Turganik settlement) may be dated to 4237–3790 cal yr BC, that is, somewhat earlier than the Holocene optimum suggested by palynologists.
Layer V shows another interval marked by increasing climate aridity and the dominance of grass steppes. As stated by the above-cited authors, the climate at the time that layer V was functioning was even dryer than during the formation of layer VI. That is confirmed by our data on the layer V composition, was formed during early Pit-Grave culture (the Early Bronze Age), in the range from 3800–3360 BC, according to the dates obtained on archeological materials (Morgunova et al., 2016b). As follows from the above, the maximum of aridity coincided with the Atlantic optimum.
It follows from the above that the Atlantic period of the Holocene was mostly characterized by arid environments; the peak of aridity fell on the early Bronze Age, the time of the early (Repino) stage of the Yamnaya culture in the Cis-Ural steppes. The Subboreal and Subatlantic periods were relatively colder and more humid, though short episodes of aridity could occur and some of them happened to be recorded in the sequence under study.
The reconstructed history of the climate changes in the Cis-Ural steppes during three intervals of the Holocene is in a good agreement with the results obtained in other regions. According to Alexandrovskiy (1996, 2000; Alexandrovskiy et al., 1999, 2004), the Atlantic period was the most arid one in the south of Russia, the subsequent intervals being comparatively wetter and colder. The extreme aridity was recorded on the Ukraine territory at the final Atlantic period, a few less arid chrono-intervals having been identified over the entire period (Kotova, 2009).
There are, however, other schemes of climate fluctuations in the central part of the Russian steppe zone; a few of them consider the Atlantic period to be humid, or even the most humid, as compared with the second half of the Holocene (Ivanov, 1992; Demkin, 1997). Also acceptable is a scenario of climatic fluctuations occurring at different times in different regions (Chendev et al., 2010). Further investigations and accumulation of empirical data would help to gain a better insight into the problem.
Conclusions
The ancient people inhabited the place from 5000 to 4000 BC (actually throughout the Atlantic period), when the place was not subjected to flooding. At the time of human habitation, the climate was mostly arid. Paleosols of that time are attributable to the Kastanozems (Endosalic Protosodic). They developed under grass (or herb and grass) steppes. The peak of aridity falls on the final Atlantic period. At the end of Eneolithic epoch (the fifth millennium BC) and in the Early Bronze Age (the fourth millennium BC) there were short-term but violent floods, which forced people to leave the habitable place.
During the Subboreal and Subatlantic periods of the Holocene, the climate became more humid, the floods became regular, the vegetation was dominated by meadow forbs and herbs growing on meadow-chernozem soils (Luvic Chernozem [Stagnic]), and the settlement was completely abandoned. In general, the studied sedimentary record at the Turganik archeological site reveals traceable climate change towards lower temperatures and increasing humidity in the second part of the Holocene, with occasional episodes of aridity that did not affect the general trend.
Interesting also the paper Collagen stable isotopes provide insights into the end of the mammoth steppe in the central East European plains during the Epigravettian, by Drucker et al., Quaternary Research (2018), 1-13
About the studied site
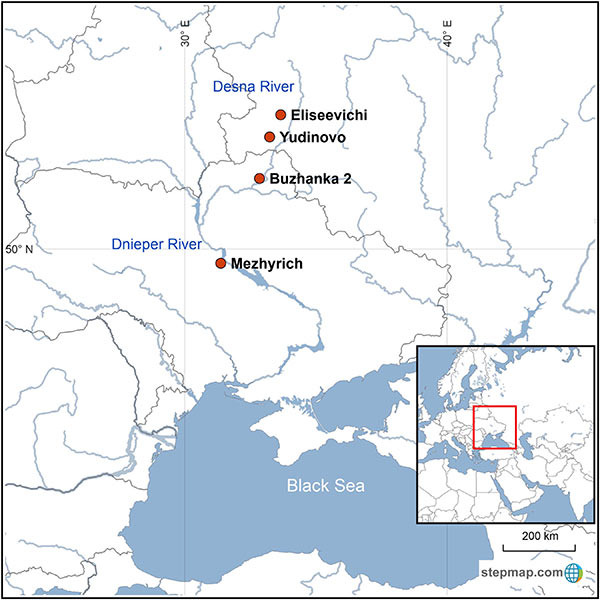
The central East European plains are famous for their Epigravettian sites that date to around 15–12 14C ka BP (ca. 18.2–13.8 cal ka BP) and display impressive large structures made from the bones of woolly mammoth (Mammuthus primigenius; e.g., Gladkih et al., 1984; Soffer, 1985; Hoffecker, 2002). The origin of the large accumulations of mammoth remains is still a matter of debate, with the main hypotheses being the collection of natural occurrences versus active hunting (e.g., Soffer, 1985; Haynes 1989; Svoboda et al., 2005). In favor of this second scenario, studies of the mammoth remains of Yudinovo (Germonpré et al., 2008) concluded that the mammoths were hunted and, at Mezhyrich, mammoths were obtained by combined procurement via collection of carcasses and active hunting (Péan, 2015). Hunting practices were observed in older sites of the Gravettian culture in the Dnieper and Desna valleys (Demay et al., 2016).
Between ca. 22 and 12 14C ka BP (ca. 26.2–13.8 ka cal BP), the Dnieper and Desna basins correspond to the southern part of the geographical distribution of the woolly mammoth (Markova et al., 2013; Kahlke, 2014). Over time their distribution shifted northwards, while the density of the mammoth population decreased (Markova et al., 2013). According to Markova et al. (2013), the combined effect of gradual warming and growing human pressure is most likely to have had a negative impact on the mammoths, resulting in their local extinction in the Russian and Ukrainian plains around 14–12 14C kaBP (ca. 17.0–13.8 ka cal BP; Stuart et al., 2005).
Discussion and conclusion
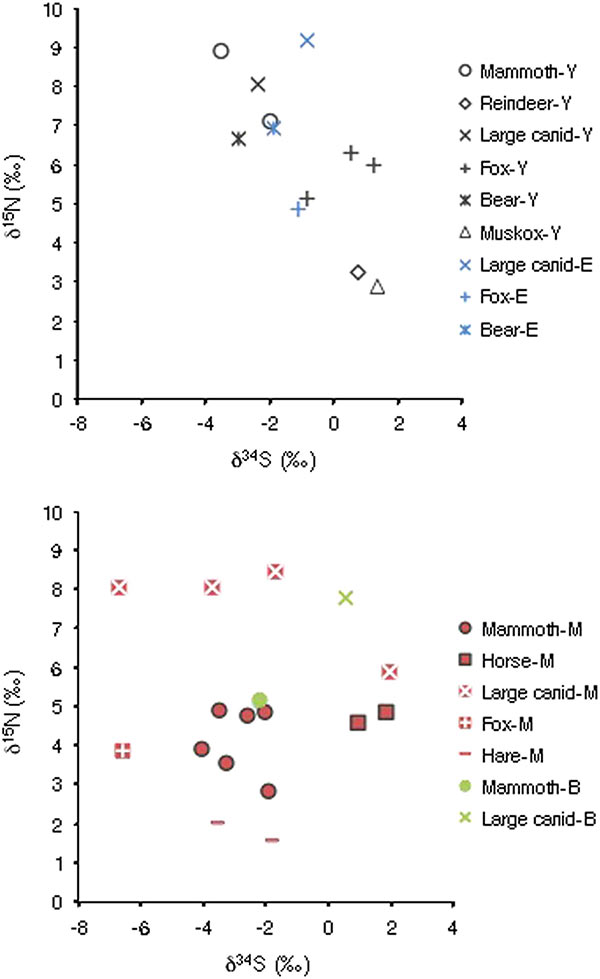
Humans could have taken advantage of the mammoth vulnerability, as reflected by lower δ15N values, to access animals that died naturally, collect bones, and hunt the most fragile individuals. This, with the possible assistance of domesticated dogs as hunting partners, could have countered a possible return to more suitable conditions for mammoth (Sablin and Khlopachev, 2002; Shipman, 2015). Our results confirm at least that mammoth specimens from Mezhyrich, Buzhanka 2, and, to a lesser extent, Eliseevichi were part of late mammoth populations surviving in sub-optimal conditions. They were thus most likely vulnerable to any pressure from environmental and/or human origin. Detecting further such cases among the late surviving mammoth populations using stable isotopic tracking may be a way to test if mammoth populations still had an optimal ecology or were metastable and, therefore, vulnerable to extinction. For instance, the insular Holocene population of mammoth on Saint Paul Island exhibited low and variable δ15N values, indicating suboptimal ecological conditions preceding their final disappearance (Graham et al., 2016).
The results of δ34S analyses showed no differences among mammoth according to the site but possibly a forage range partitioning between mammoth and coexisting large ungulates. Thus, variability in the mobility pattern for the mammoth between the high and low δ15N groups, such as migratory versus sedentary individuals, is not supported so far. We consider that rapid environmental modifications over time, probably not detectable through radiocarbon dating, can be a valid alternative explanation. Combined with direct competition with other large herbivores, such as the horse, and hunting of the most vulnerable individuals, the loss of their optimal habitat was likely to be the driving factor behind the local extinction of the mammoth in the central East European plains.
Related:
- Climatic conditions in the Cis-Ural Steppe region and the Repin culture
- Steppe and Caucasus Eneolithic: the new keystones of the EHG-CHG-ANE ancestry in steppe groups
- The Caucasus a genetic and cultural barrier; Yamna dominated by R1b-M269; Yamna settlers in Hungary cluster with Yamna
- Differing modes of animal exploitation in North Pontic Eneolithic and Bronze Age Societies
- Consequences of Damgaard et al. 2018 (I): EHG ancestry in Maykop samples, and the potential Anatolian expansion routes
- On the potential origin of Caucasus hunter-gatherer ancestry in Eneolithic steppe cultures
- Eneolithic Ukraine cultures of the North Pontic steppe and southern steppe-forest, on the Left Bank of the Dnieper
- North Pontic steppe Eneolithic cultures, and an alternative Indo-Slavonic model
- No large-scale steppe migration into Anatolia; early Yamna migrations and MLBA brought LPIE dialects in Asia
- Proto-Indo-European homeland south of the Caucasus?