New paper (behind paywall) Reconstructing the genetic history of late Neanderthals, by Mateja Hajdinjak, Qiaomei Fu, Alexander Hübner, et al. Nature (2018).
Abstract (edited):
Although it has previously been shown that Neanderthals contributed DNA to modern humans, not much is known about the genetic diversity of Neanderthals or the relationship between late Neanderthal populations at the time at which their last interactions with early modern humans occurred and before they eventually disappeared. Our ability to retrieve DNA from a larger number of Neanderthal individuals has been limited by poor preservation of endogenous DNA and contamination of Neanderthal skeletal remains by large amounts of microbial and present-day human DNA. Here we use hypochlorite treatment6 of as little as 9 mg of bone or tooth powder to generate between 1- and 2.7-fold genomic coverage of five Neanderthals who lived around 39,000 to 47,000 years ago (that is, late Neanderthals), thereby doubling the number of Neanderthals for which genome sequences are available. Genetic similarity among late Neanderthals is well predicted by their geographical location, and comparison to the genome of an older Neanderthal from the Caucasus indicates that a population turnover is likely to have occurred, either in the Caucasus or throughout Europe, towards the end of Neanderthal history. We find that the bulk of Neanderthal gene flow into early modern humans originated from one or more source populations that diverged from the Neanderthals that were studied here at least 70,000 years ago, but after they split from a previously sequenced Neanderthal from Siberia around 150,000 years ago. Although four of the Neanderthals studied here post-date the putative arrival of early modern humans into Europe, we do not detect any recent gene flow from early modern humans in their ancestry.
Interesting excerpts (edited):
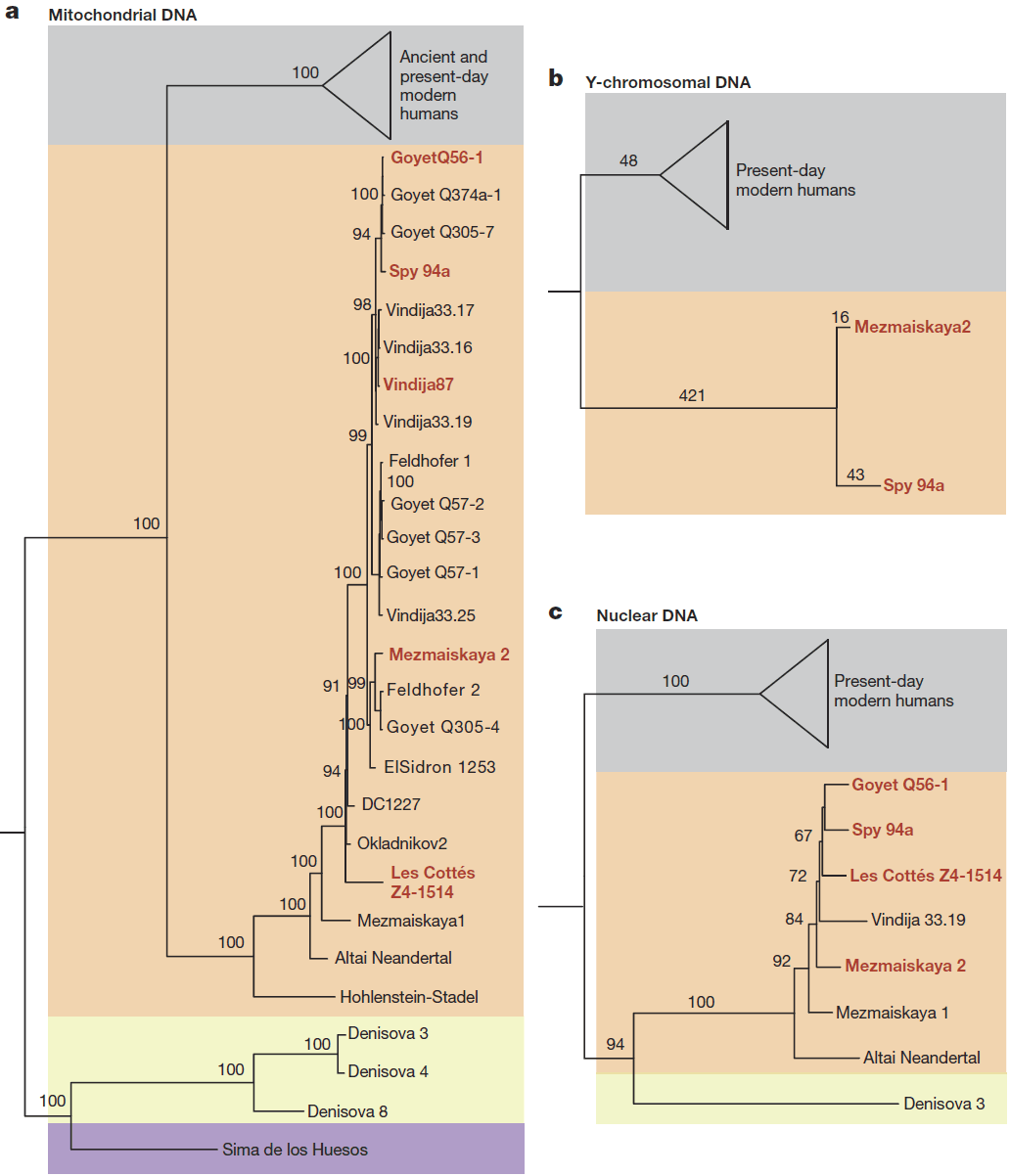
(…) Mezmaiskaya 2 shared more derived alleles with the other late Neanderthals than with Mezmaiskaya 1 (− 2.13 ≤ Z ≤ − 9.56; Supplementary Information 9), suggesting that there was a population turnover towards the end of Neanderthal history. This turnover may have been the result of a population related to western Neanderthals replacing earlier Neanderthals in the Caucasus, or the replacement of Neanderthals in western Europe by a population related to Mezmaiskaya 2. The timing of this turnover coincides with pronounced climatic fluctuations during Marine Isotope Stage 3 between 60 and 24 ka, when extreme cold periods in northern Europe may have triggered the local extinction of Neanderthal populations and subsequent re-colonization from refugia in southern Europe or western Asia.
(…) the majority of gene flow into early modern humans appears to have originated from one or more Neanderthal populations that diverged from other late Neanderthals after their split from the Altai Neanderthal about 150 ka, but before the split from Mezmaiskaya 1 at least 90 ka. Owing to the scarcity of overlapping genetic data from Oase 1, whose genome revealed an unusually high percentage of Neanderthal ancestry11, we were unable to resolve whether one of these late Neanderthals was significantly closer than others to the introgressing Neanderthal in Oase 1.
Interbreeding between Neanderthals and early modern humans is likely to have occurred intermittently, presumably resulting in gene flow in both directions. However, when we applied an approach that uses the extended length of haplotypes expected from recent introgression into the analysed late Neanderthals, we did not find any indications of recent gene flow from early modern humans to the late Neanderthals. We caution that given the small number of analysed Neanderthals we cannot exclude that such gene flow occurred. However, it is striking that Oase 1, one of two early modern humans that overlapped in time with late Neanderthals, showed evidence for recent additional Neanderthal introgression whereas none of the late Neanderthals analysed here do. This may indicate that gene flow affected the ancestry of modern human populations more than it did Neanderthals
Related:
- Two sources of archaic Denisovan ancestry in East Asia, one possibly after the isolation of Native Americans
- Review article about Ancient Genomics, by Pontus Skoglund and Iain Mathieson
- Review article on the origin of modern humans: the multiple-dispersal model and Late Pleistocene Asia
- Ancient genomes document multiple waves of migration in south-east Asian prehistory
- Genomics reveals four prehistoric migration waves into South-East Asia
- Population turnover in Remote Oceania shortly after initial settlement
- Genomic history of South-East Asia: eastern Polynesians, Peninsular Malaysia and North Borneo
- Islands across the Indonesian archipelago show complex patterns of admixture
- Two more studies on the genetic history of East Asia: Han Chinese and Thailand
- Reconstructing the demographic history of the Himalayan and adjoining populations
- Ancient Di-Qiang people show early links with Han Chinese