Wang et al. (2018) is obviously a game changer in many aspects. I have already written about the upcoming Yamna Hungary samples, about the new Steppe_Eneolithic and Caucasus Eneolithic keystones, and about the upcoming Greece Neolithic samples with steppe ancestry.
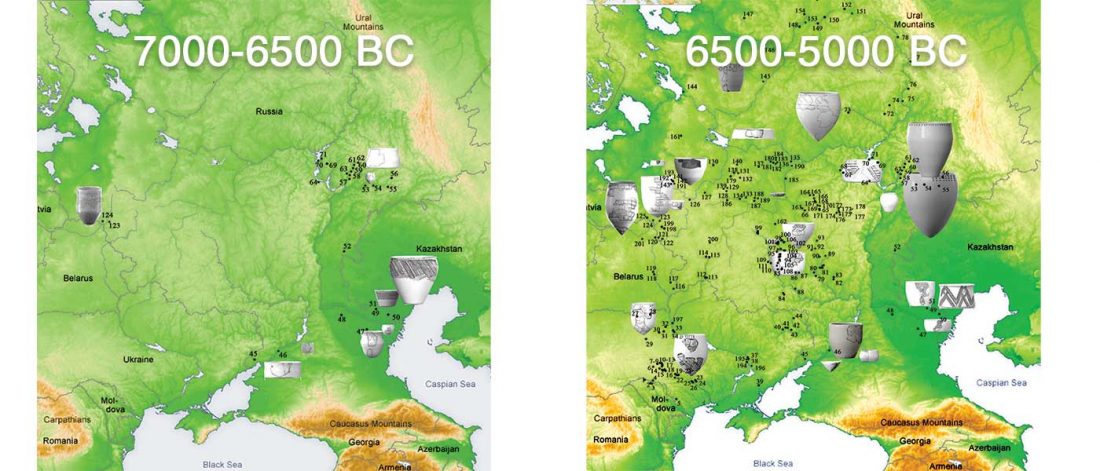
An interesting aspect of the paper, hidden among so many relevant details, is a clearer picture of how the so-called Yamnaya or steppe ancestry evolved from Samara hunter-gatherers to Yamna nomadic pastoralists, and how this ancestry appeared among Proto-Corded Ware populations.
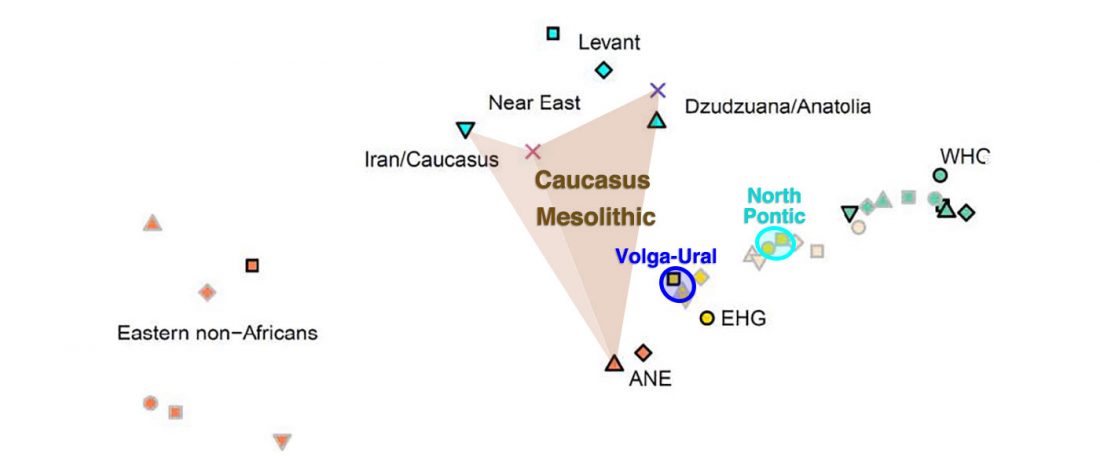
Please note: arrows of “ancestry movement” in the following PCAs do not necessarily represent physical … Read the rest ““Steppe ancestry” step by step: Khvalynsk, Sredni Stog, Repin, Yamna, Corded Ware”