Open access research article Decline of genetic diversity in ancient domestic stallions in Europe, by Wutke et al., Science (2018), 4(4):eaap9691.
Abstract (emphasis mine):
Present-day domestic horses are immensely diverse in their maternally inherited mitochondrial DNA, yet they show very little variation on their paternally inherited Y chromosome. Although it has recently been shown that Y chromosomal diversity in domestic horses was higher at least until the Iron Age, when and why this diversity disappeared remain controversial questions. We genotyped 16 recently discovered Y chromosomal single-nucleotide polymorphisms in 96 ancient Eurasian stallions spanning the early domestication stages (Copper and Bronze Age) to the Middle Ages. Using this Y chromosomal time series, which covers nearly the entire history of horse domestication, we reveal how Y chromosomal diversity changed over time. Our results also show that the lack of multiple stallion lineages in the extant domestic population is caused by neither a founder effect nor random demographic effects but instead is the result of artificial selection—initially during the Iron Age by nomadic people from the Eurasian steppes and later during the Roman period. Moreover, the modern domestic haplotype probably derived from another, already advantageous, haplotype, most likely after the beginning of the domestication. In line with recent findings indicating that the Przewalski and domestic horse lineages remained connected by gene flow after they diverged about 45,000 years ago, we present evidence for Y chromosomal introgression of Przewalski horses into the gene pool of European domestic horses at least until medieval times.
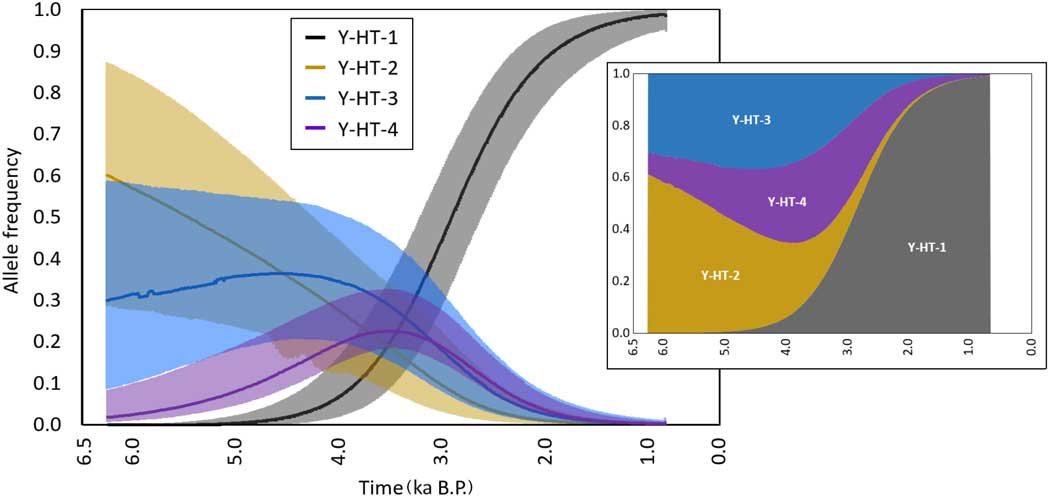
Inferred temporal trajectories of haplotype frequencies. Each haplotype is displayed by a different color. The shaded area represents the 95% highest-density region. The trajectories were constructed taking the median values across frequencies from the simulations of the Bayesian posterior sample. The small chart represents the stacked frequencies; the amplitude of each colored area is proportional to the median haplotype frequencies (normalized) at a given time. The x and y axes of the small chart match those in the large one. Ka, thousands of years.
Interesting excerpts:
The first record of the modern domestic Y chromosome haplotype stems from two Bronze Age samples of similar age. Notably, both samples were found in two distantly located regions: present-day Slovakia (2000–1600 BCE, dated by archaeological context) and western Siberia (14C-dated: 1609–1436 cal. BCE). Although a very recent study proposes an oriental origin of this haplotype (14), we cannot determine the geographical origin of Y-HT-1 with certainty, because this haplotype has not been found thus far in predomestic or wild stallions. There are two possible scenarios: (i) Y-HT-1 emerged within the domestic population by mutation and (ii) Y-HT-1 was already present in wild horses and entered the domestic population either at the beginning of domestication (but initially restricted to Asian horses) or later by introgression (from wild Y-HT-1 carrying studs during the Iron Age). Crosses between domestic animals and their wild counterparts have been observed in several domestic species (15–18); thus, the simplest explanation would be that we missed Y-HT-1 in older samples because of limited geographical sampling. However, the estimated haplotype age is contemporary (Fig. 4) with the assumed starting point of horse domestication ~4000–3500 BCE (19), rendering it likely that Y-HT-1 originated within the domestic horse gene pool. Still, we cannot rule out definitively that it appeared before domestication.
Independent of its geographical origin, Y-HT-1 progressively replaced all other haplotypes—except for one additional lineage that is restricted to Yakutian horses (11). Considering our data, this trend in paternal diversity toward dominance of the modern lineage appears to start in the Bronze Age and becomes even more pronounced during the Iron Age. The Bronze Age was a time of large-scale human migrations across Eurasia (20–22), movements that were undoubtedly facilitated by the spread of horses as a means of transport and warfare. At that time, the western Eurasian steppes were inhabited by highly mobile cultures that largely relied on horses (20, 21, 23, 24). The genetic admixture of northern and central European humans with Caucasians/eastern Europeans did correlate with the spread of the Yamnaya culture from the Pontic-Caspian steppe (25), an area that has repeatedly been suggested as the center of horse domestication (19, 26, 27). Given the importance of domestic horses, it appears that deliberate selection/rejection of certain stallions by these people might have contributed to the loss of paternal diversity. The spread of humans out of this region might also have resulted in the spread of Y-HT-1 from Asia to Europe. This scenario also agrees with recent findings that the low male diversity of extant horses is not caused by recruiting only a limited number of stallions during early domestication (13).
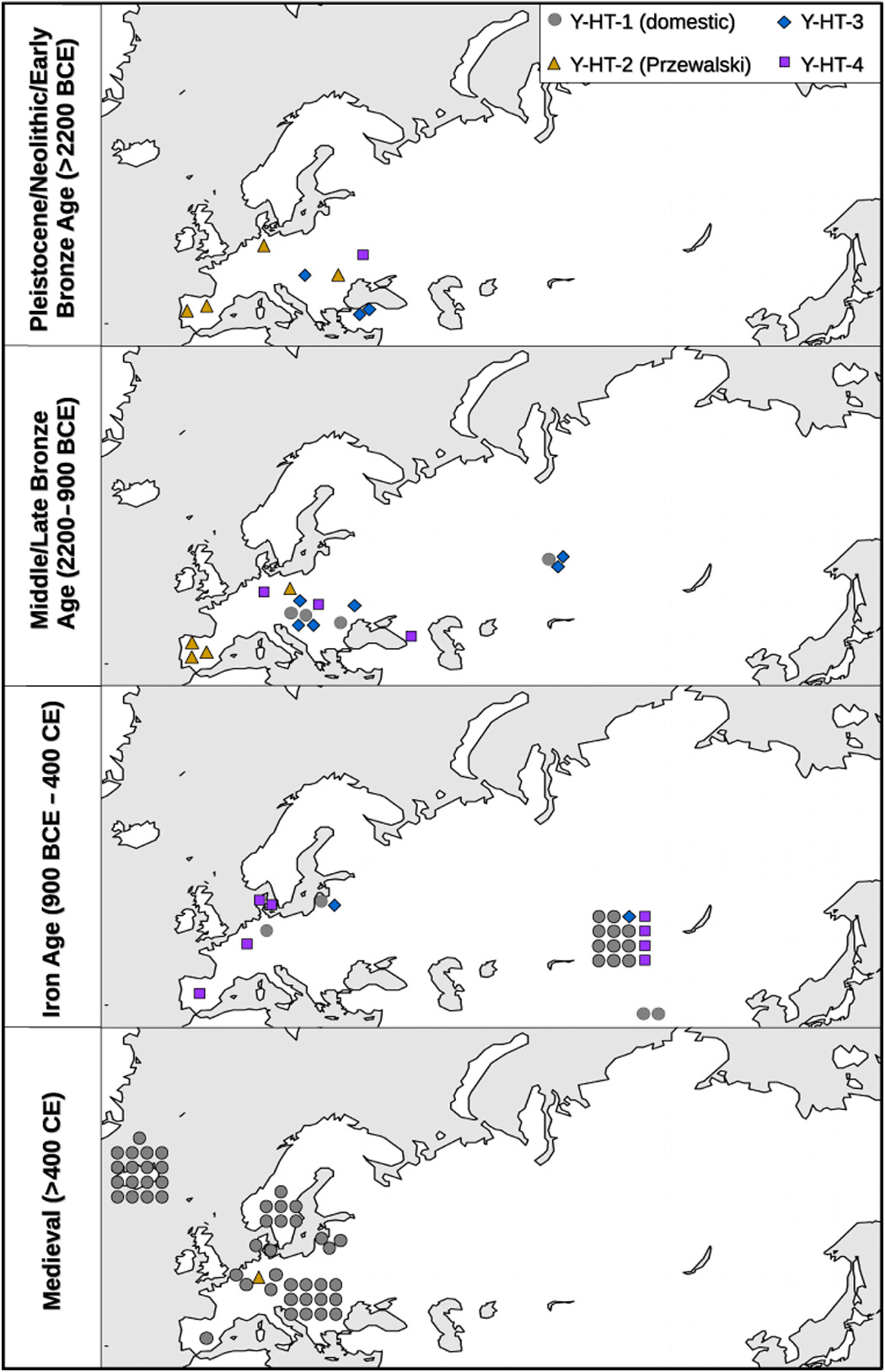
Maps displaying age, locality, and haplotype (different colors) of each successfully genotyped sample.
The presence of the Y chromosome haplotype carried by present-day Przewalski horses (Y-HT-2) in early domestic stallions and a European wild horse (Pie05; table S2) could be the result of introgression of Przewalski stallions. Although the original distribution of the Przewalski horse is unknown, it was probably much larger than that of the relict population in Mongolia that produced modern Przewalski horses and might even have extended into Central Europe. However, it is also possible that either Przewalski horses were among the initially domesticated horses or that Y-HT-2 occurred both in Przewalski horses and in those wild horses that are the ancestors of domestic horses, based on autosomal DNA data (30). Regardless of how Y-HT-2 entered the domestic gene pool, it was eventually lost, as were all haplotypes except Y-HT-1. In our sample set, Y-HT-2 was undetectable as early as the third time bin. However, it is possible that Y-HT-2 may have been present during this time period, but with a frequency below 0.11 (with 95% probability). The inferred time trajectories for Y-HT-2 frequencies suggest that it could nevertheless have persisted at very low frequencies until the Middle Ages (Fig. 3). On the basis of these simulations, this finding could be interpreted as a relic of this haplotype’s formerly higher frequency in the domestic horse gene pool. It is also possible that the presence of this haplotype could be the result of mating a wild stallion with a domestic mare, a frequently reported breeding practice when wild horses were still widely distributed. However, a significant contribution of the Przewalski horse to the gene pool of modern domestic horses has been almost ruled out by recent genomic studies (13, 31, 32).
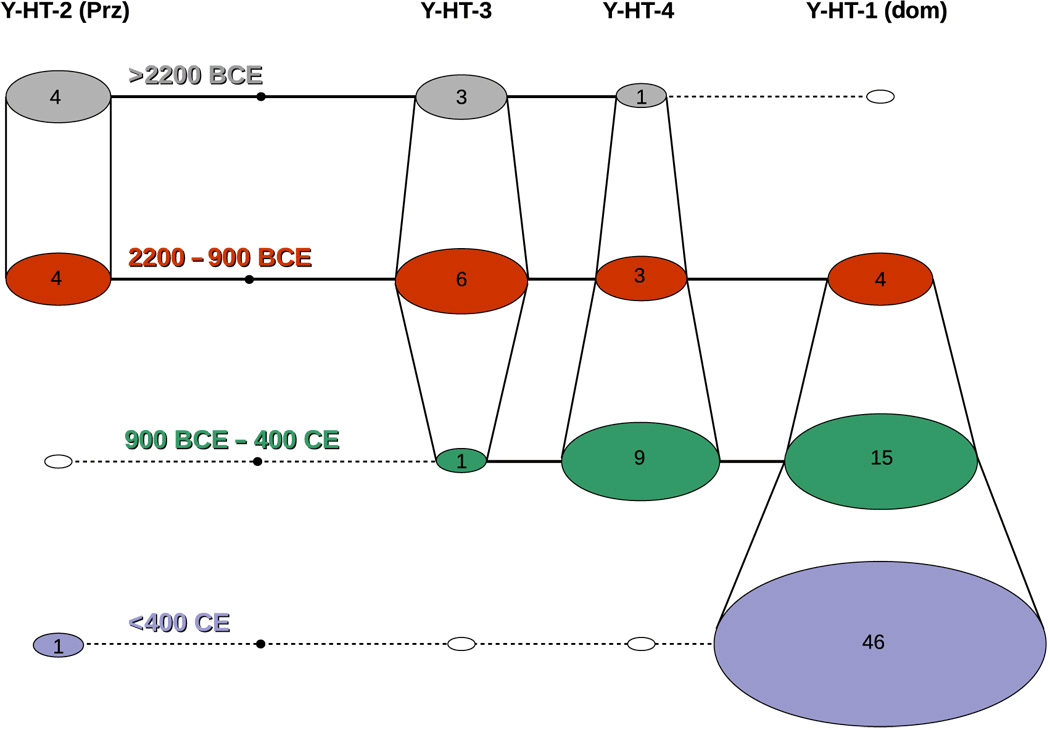
Temporal haplotype network of the four detected Y chromosome haplotypes. Age of the samples indicated by multiple layers separated by color; vertical lines connecting the haplotypes of consecutive layers/ages represent which haplotype was transferred into a later/younger period. Numbers constitute the respective number of individuals showing this particular haplotype for that period. Prz, Przewalski; Dom, domestic.
Related:
- Ancient DNA upends the horse family tree
- Proto-Indo-European homeland south of the Caucasus?
- On the potential origin of Caucasus hunter-gatherer ancestry in Eneolithic steppe cultures
- Consequences of O&M 2018 (II): The unsolved nature of Suvorovo-Novodanilovka chiefs, and the route of Proto-Anatolian expansion
- Consequences of O&M 2018 (I): The latest West Yamna “outlier”
- Olalde et al. and Mathieson et al. (Nature 2018): R1b-L23 dominates Bell Beaker and Yamna, R1a-M417 resurges in East-Central Europe during the Bronze Age
- The Indo-European demic diffusion model, and the “R1b – Indo-European” association
- The concept of “Outlier” in Human Ancestry (III): Late Neolithic samples from the Baltic region and origins of the Corded Ware culture