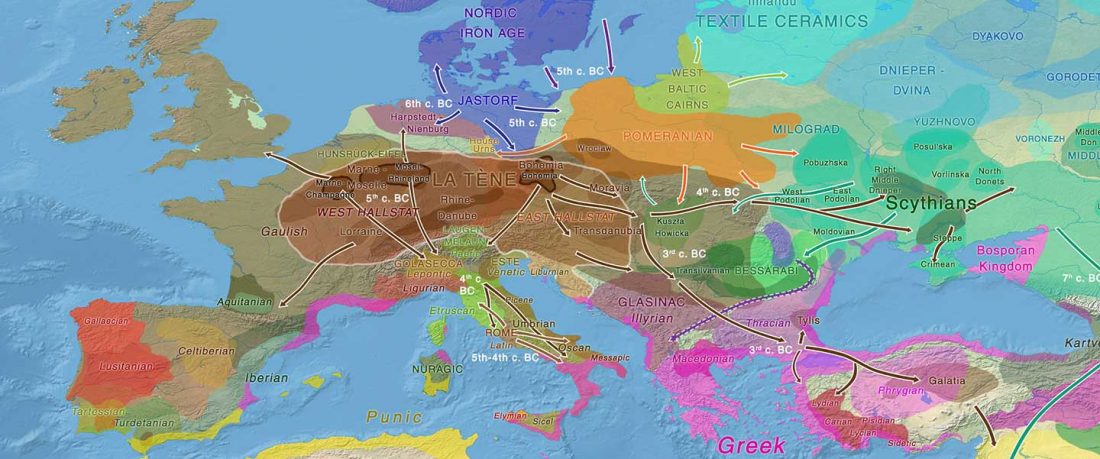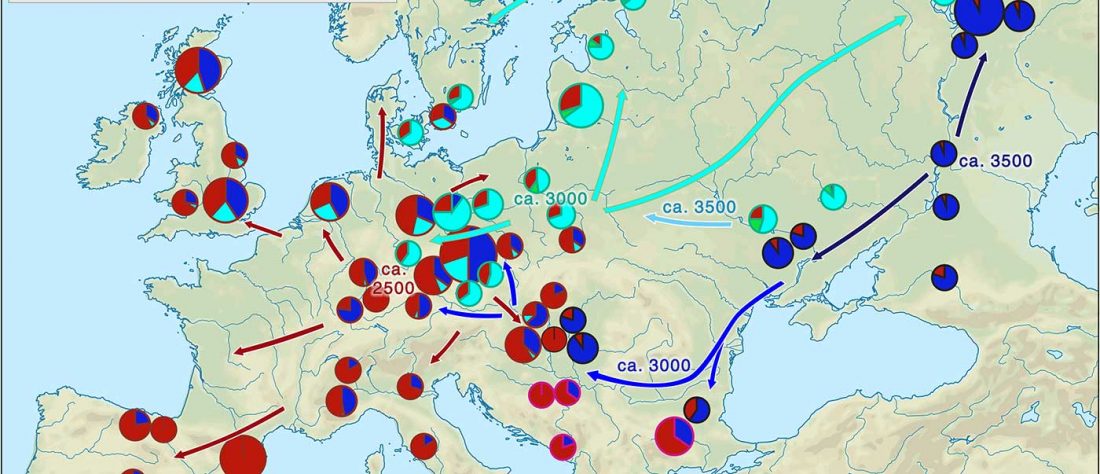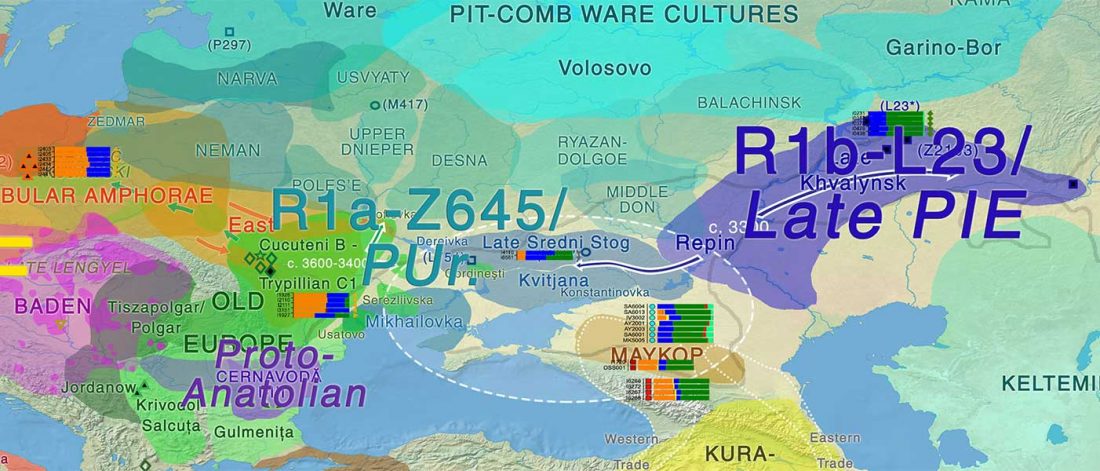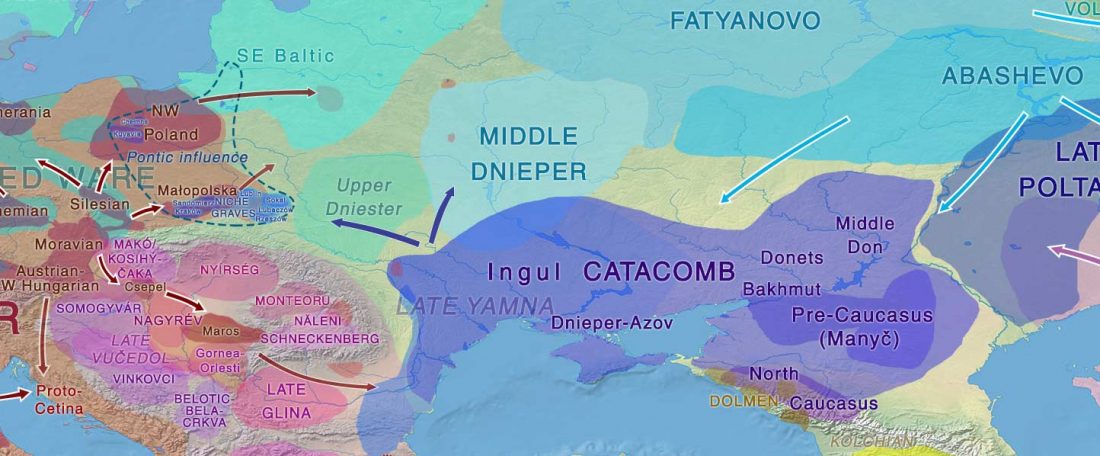
In the recent Linderholm et al. (2020), I preferred to interpret the finding of R1b-P310* among late niche (catacomb) grave groups of Lesser Poland as derived from Late PIE – Late Uralic contacts, through a much earlier intrusion of late Repin/early Yamnaya chieftains among Late Trypillians.
This is one of the few aspects of the books where I tried to offer my own contribution to the field, by combining the Indo-Uralic concept (which supports a distinct evolution of laryngeals for PIE and PU) with a modified, ‘layered’ use of Koivulehto’s controversial and irregular PIE laryngeal borrowing as PU … Read the rest “Indo-Iranian influence on West Uralic through the Catacomb culture”