Researchers involved in the investigation of the Yampil Barrow Complex are taking the opportunity of their latest genetic paper to publish and upload more papers in Academia.edu.
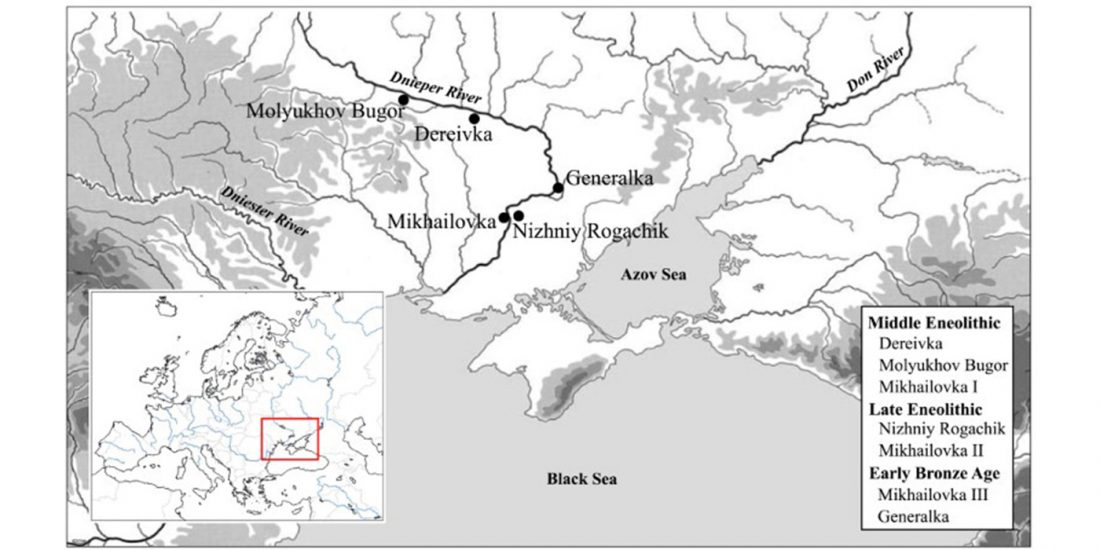
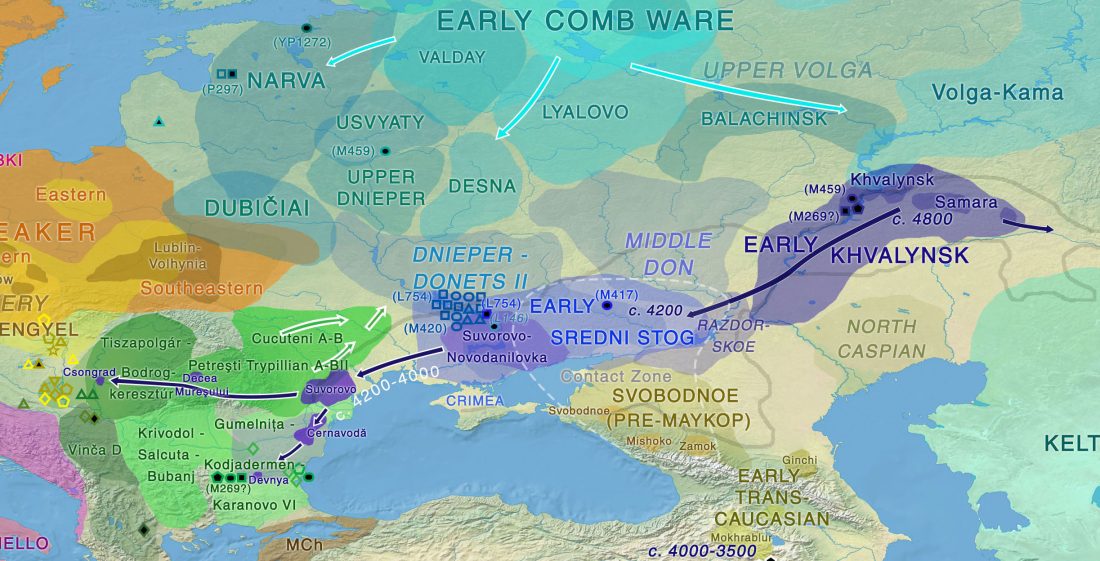
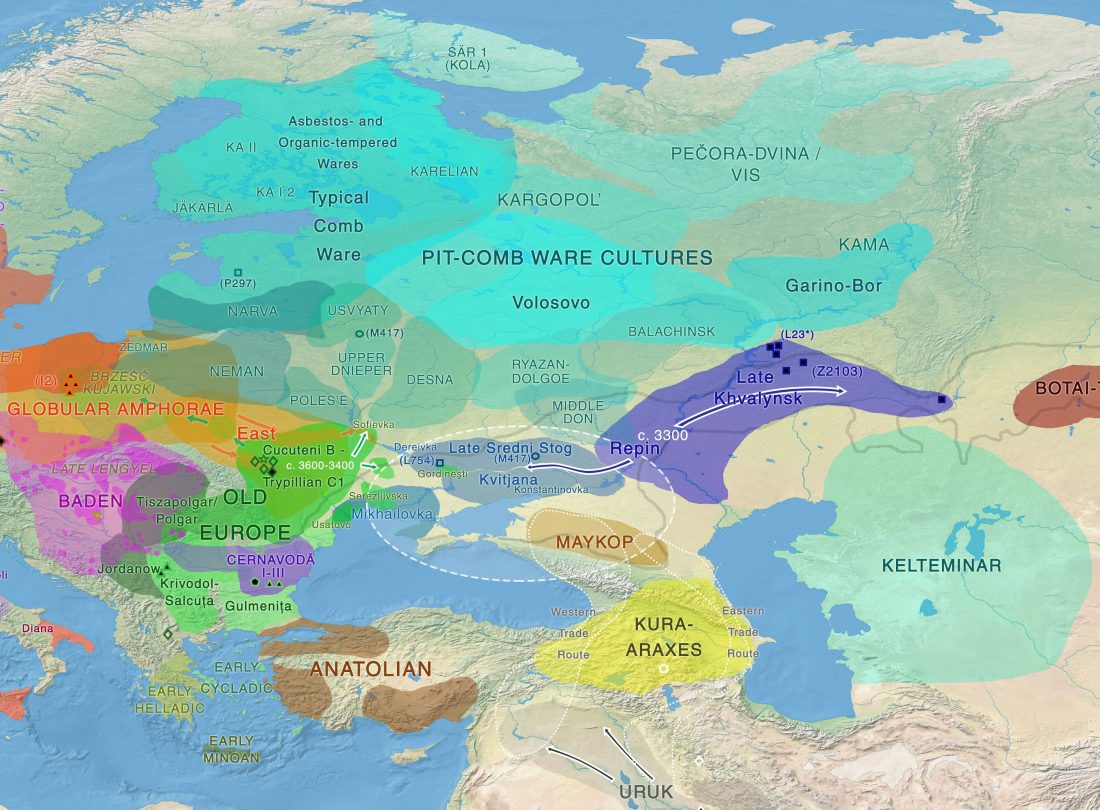
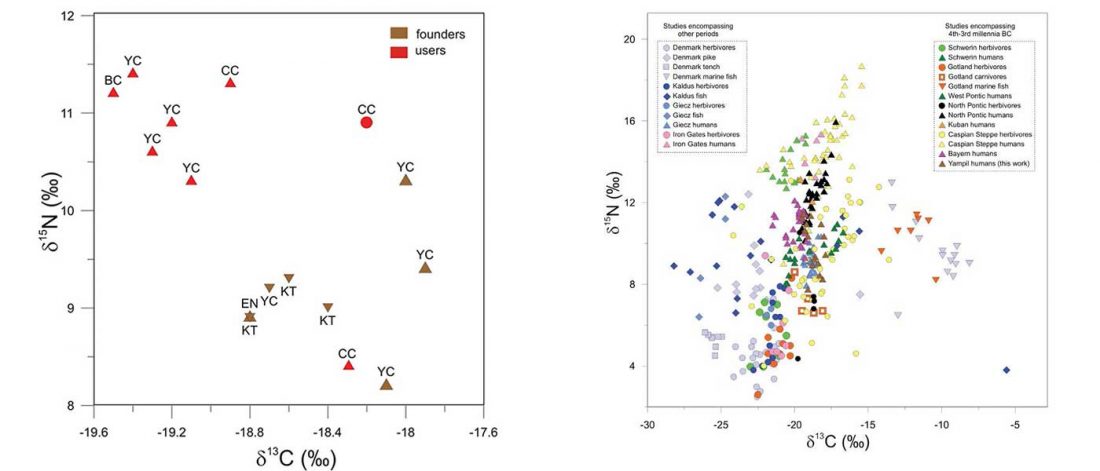
NOTE. These are from the free volume 22 of Baltic-Pontic Studies, Podolia “Barrow Culture” Communities: 4th/3rd-2nd Mill. BC. The Yampil Barrow Complex: Interdisciplinary Studies, whose website gives a warning depending on your browser (because of the lack of secure connection). Here is a link to the whole PDF.
Here are some of them, with interesting excerpts (emphasis mine):
1. Kurgan rites in the Eneolithic and Early Bronze age Podolia in … Read the rest “The Yampil Barrow complex and the Yamna connection with forest-steppe cultures”