Open access Late Quaternary horses in Eurasia in the face of climate and vegetation change, by Leonardi et al. Science Advances (2018) 4(7):eaar5589.
Interesting excerpts (emphasis mine):
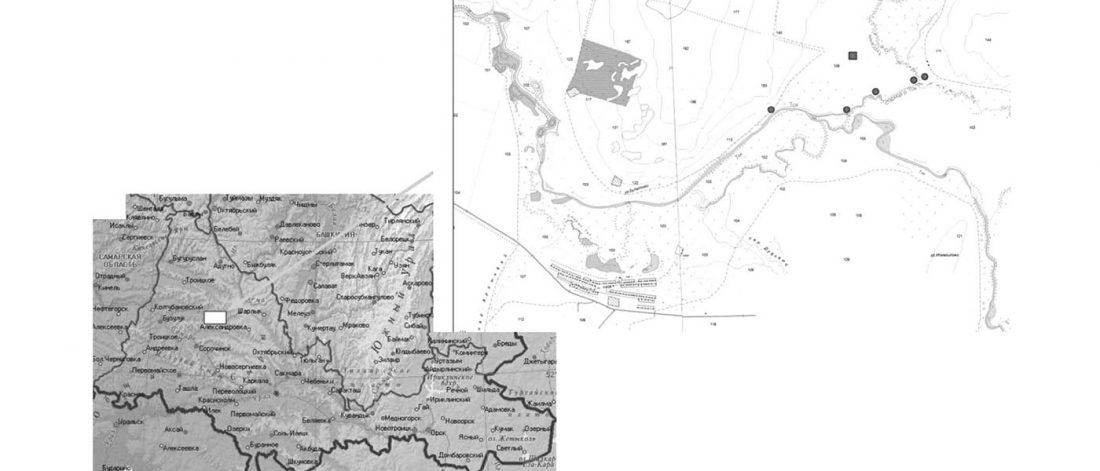
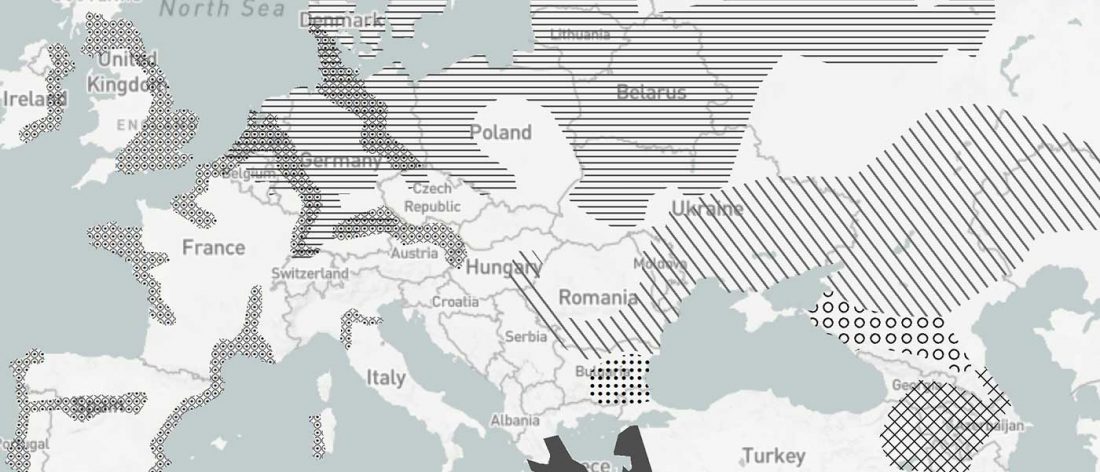
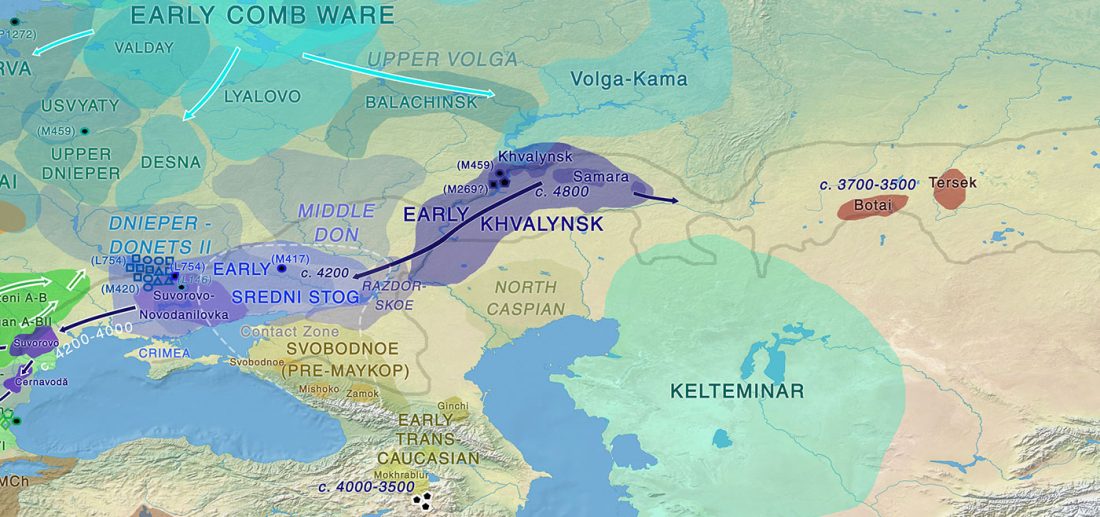
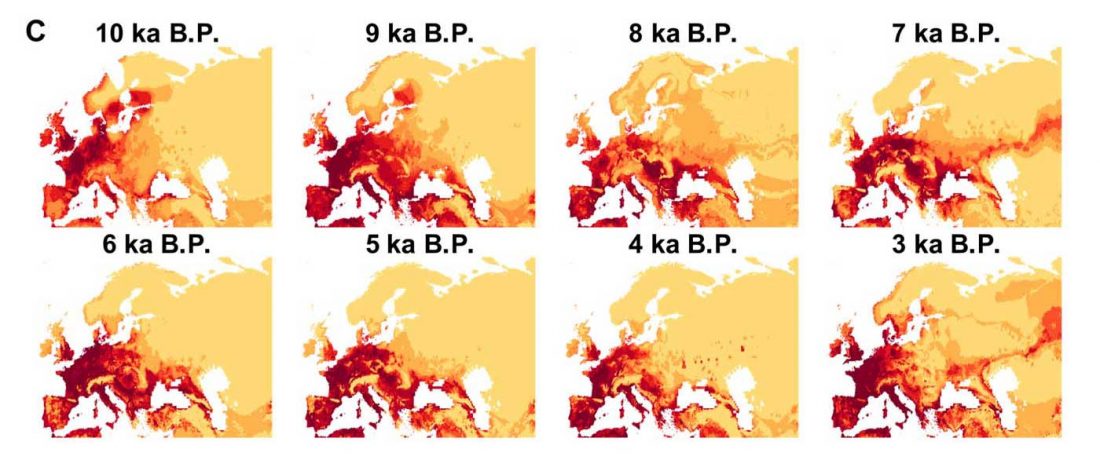
… Read the rest “Origin of horse domestication likely on the North Caspian steppes”Here, we compiled an extensive continental-scale database, consisting of 3070 radiocarbon dates associated to horse paleontological and archeological finds across the whole of Eurasia, that has been analyzed in association with coarse-scale paleoclimatic reconstructions. We further collected the number of identified specimens (NISP) frequency data for horses versus other ungulates in 1120 archeological layers in Europe (…) This ma.ssive amount of data allowed us to track,with unprecedented details, how the geographic