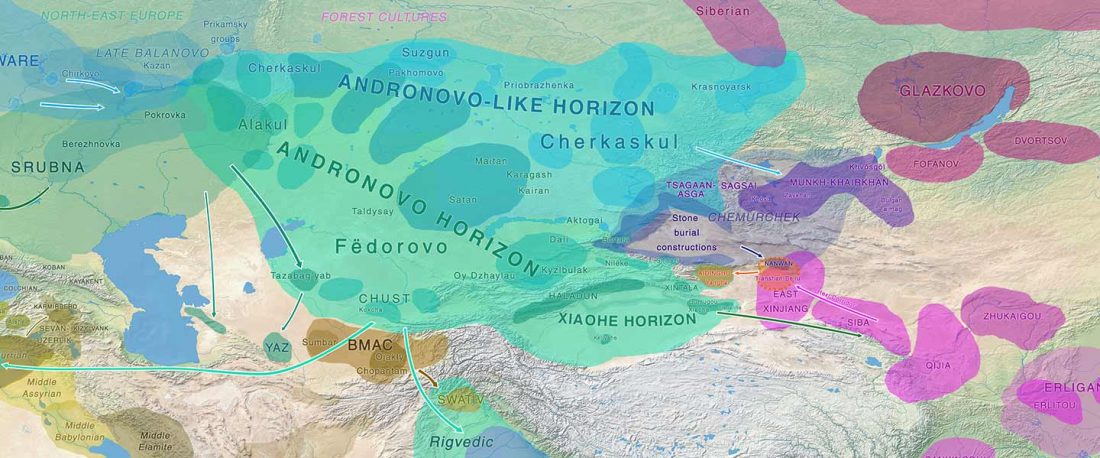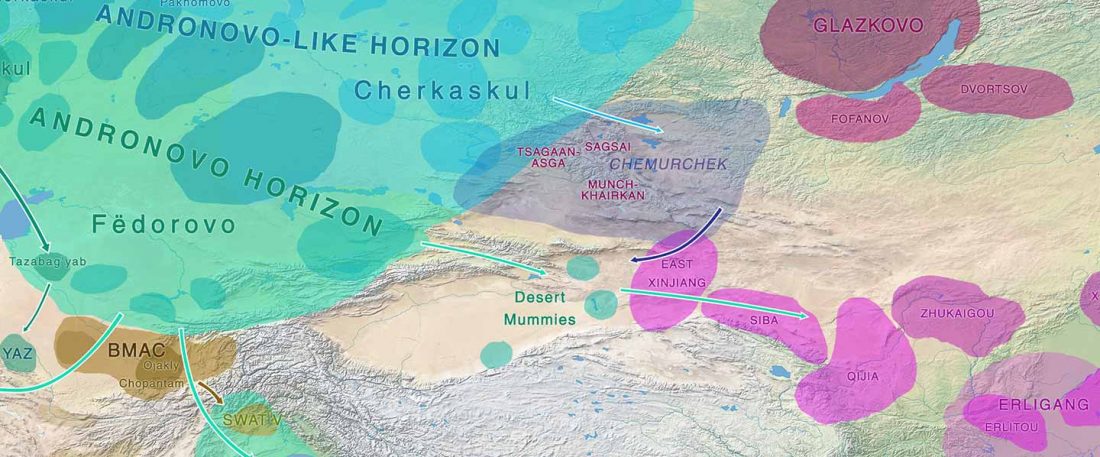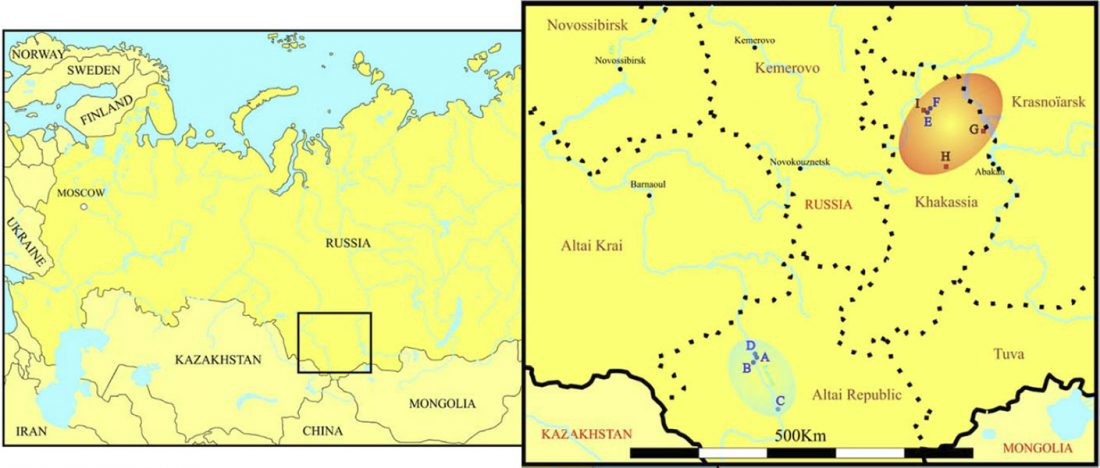
Recent paper (behind paywall) The Baigetuobie cemetery: New discovery and human genetic features of Andronovo community’s diffusion to the Eastern Tianshan Mountains (1800–1500 BC), by Zhu, Zhang, et al. The Holocene (2020).
Interesting excerpts (emphasis mine):
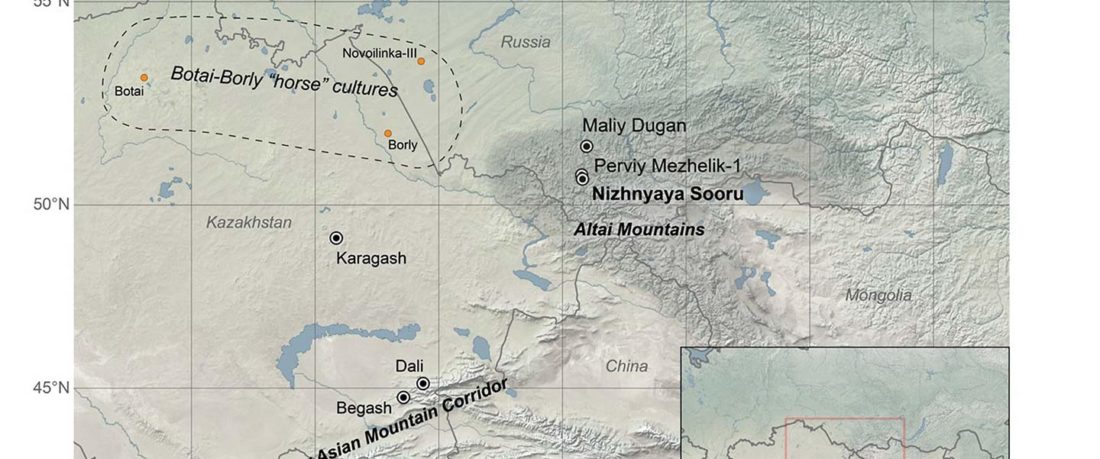
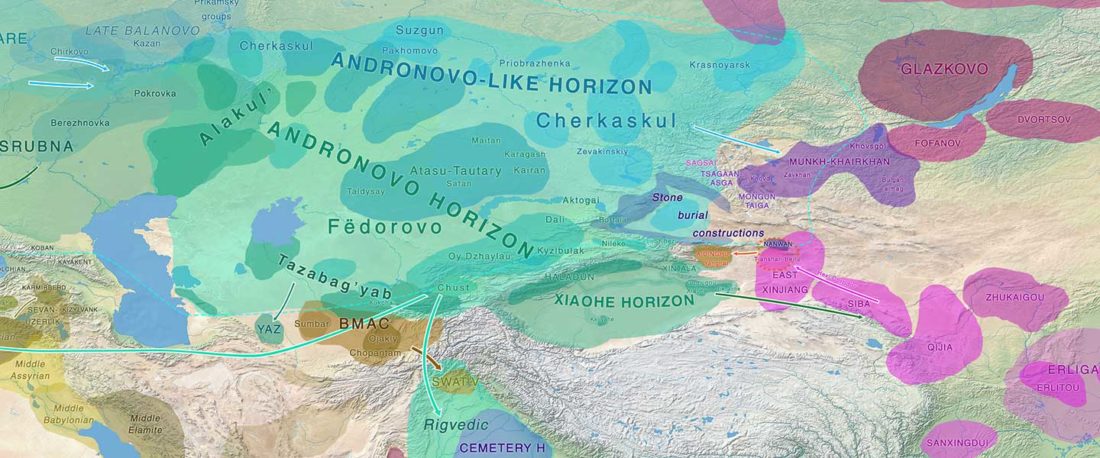
The Baigetuobie cemetery (Balikun County, Hami City) is located on the eastern slope of the Baigetuobie Mountains, about 54 km west of the Balikun County and 40 km southwest of the Balikun Lake.
Baigetuobie cemetery
… Read the rest “Early Andronovo intrusion in the Eastern Tianshan”The cemetery is relatively small in scale and the burials are rather concentrated. The graves are marked on the surface by stone slabs visible from approximately 0.05 to