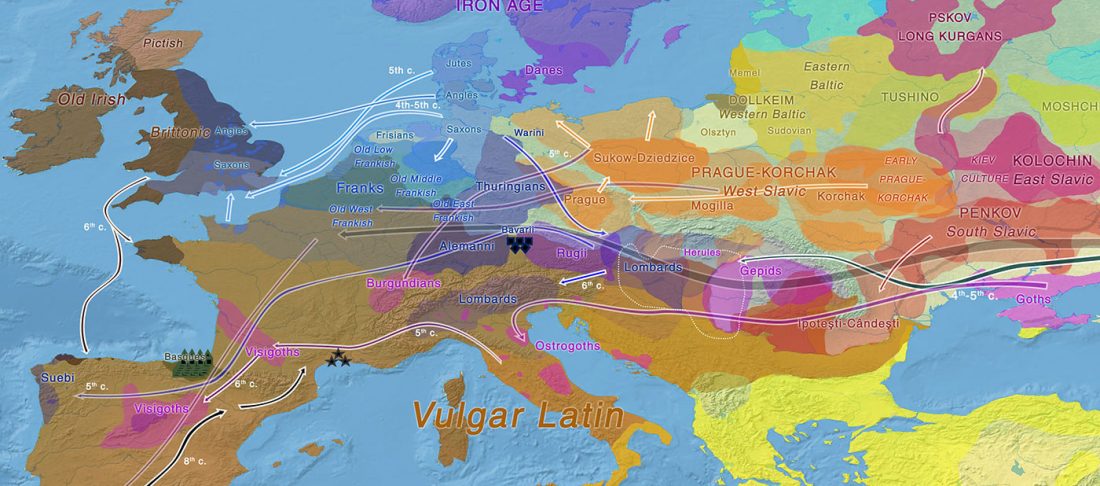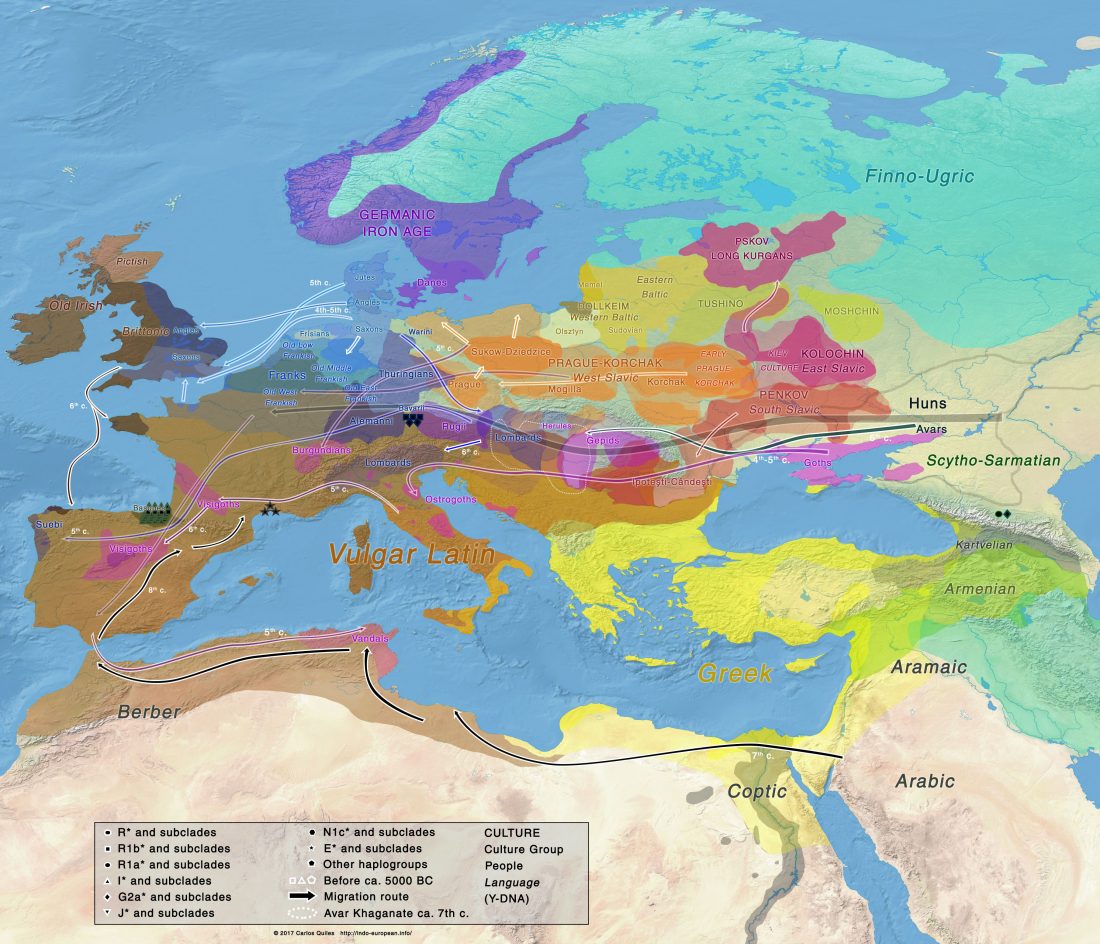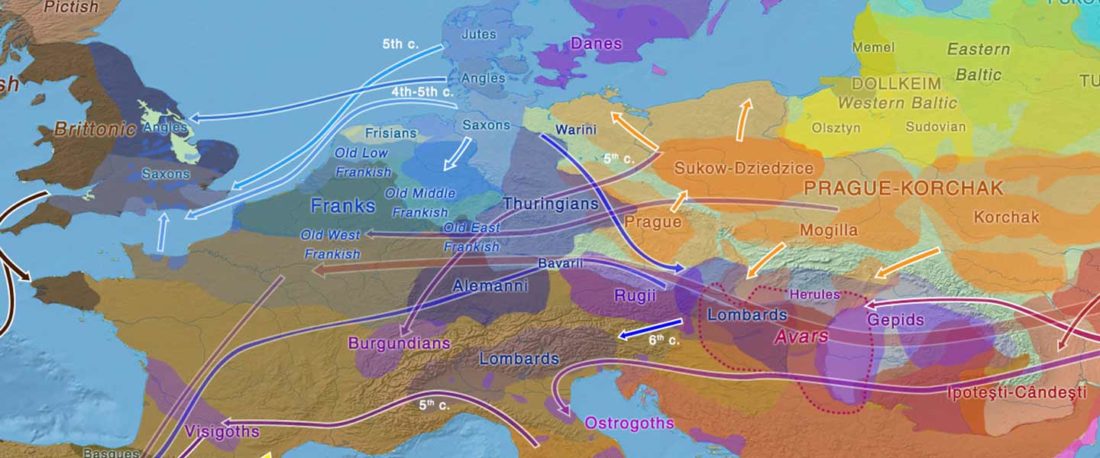
The Family Tree DNA R&D team formed by Göran Runfeldt and Michael Sager has reported detailed Y-SNPs of sampled Longobards from the open access paper Understanding 6th-century barbarian social organization and migration through paleogenomics, by Amorim et al. Nat. Commun. (2020). From the abstract:
… Read the rest “Longobards from Scandinavia, and the “Ural-Altaic” Árpád lineage”We obtained ancient genomic DNA from 63 samples from two cemeteries (from Hungary and Northern Italy) that have been previously associated with the Longobards, a barbarian people that ruled large parts of Italy for over 200 years after invading from Pannonia in 568 CE. Our dense cemetery-based sampling revealed that each cemetery was primarily organized