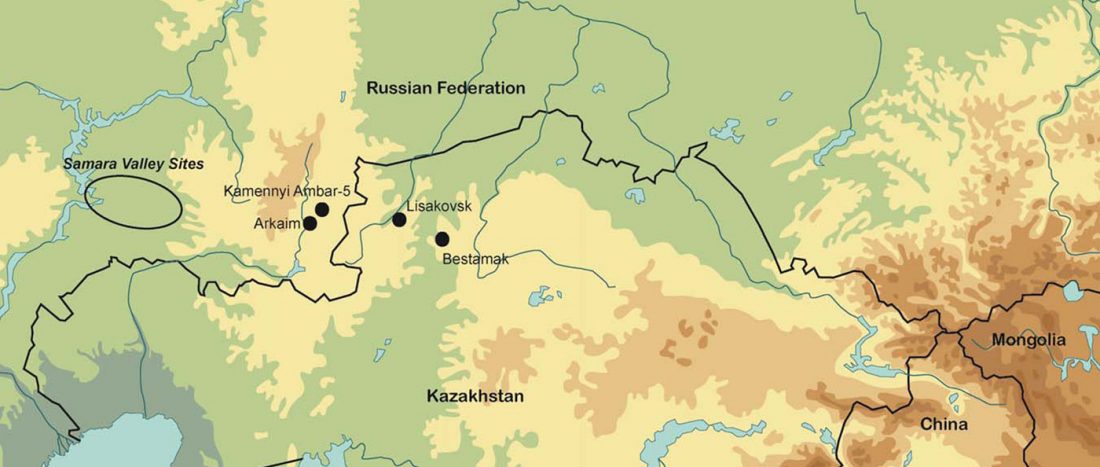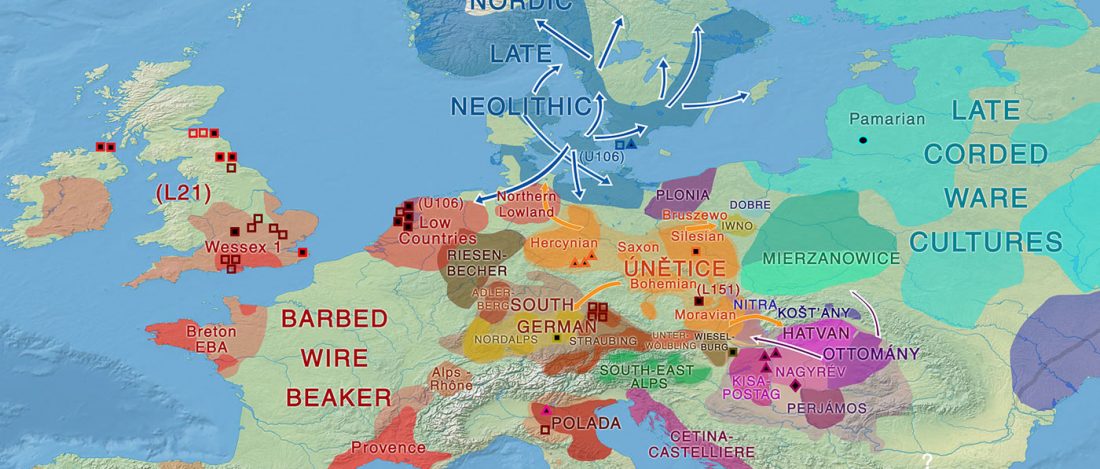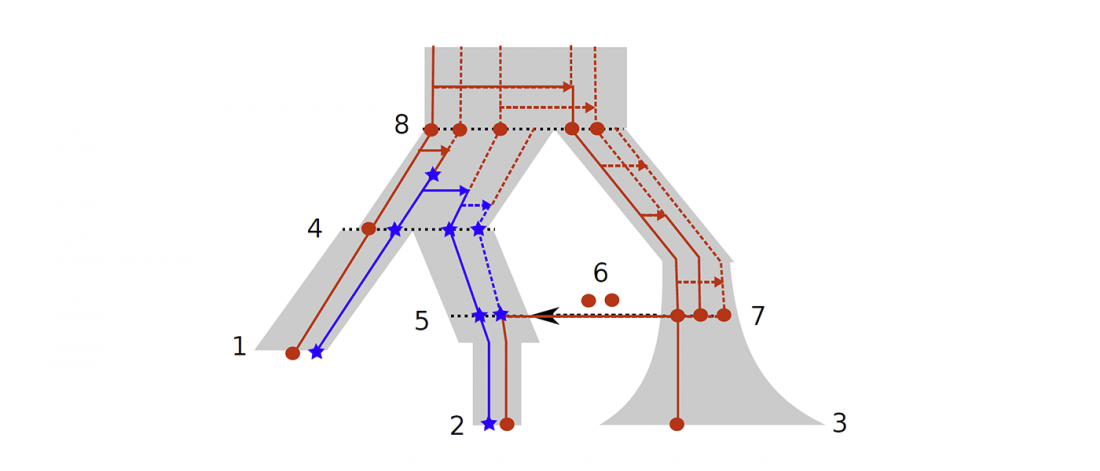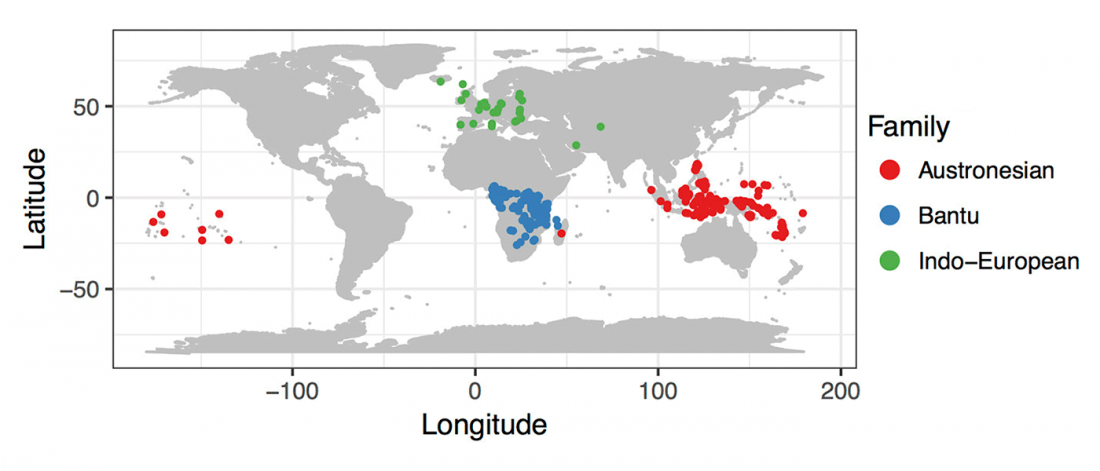
Open access Population Size and the Rate of Language Evolution: A Test Across Indo-European, Austronesian, and Bantu Languages, by Greenhill et al. Front. Psychol (2018) 9:576.
Summary (emphasis mine):
… Read the rest “Population size potentially affecting rates of language change”What role does speaker population size play in shaping rates of language evolution? There has been little consensus on the expected relationship between rates and patterns of language change and speaker population size, with some predicting faster rates of change in smaller populations, and others expecting greater change in larger populations. The growth of comparative databases has allowed population size effects to be investigated across a wide range of language