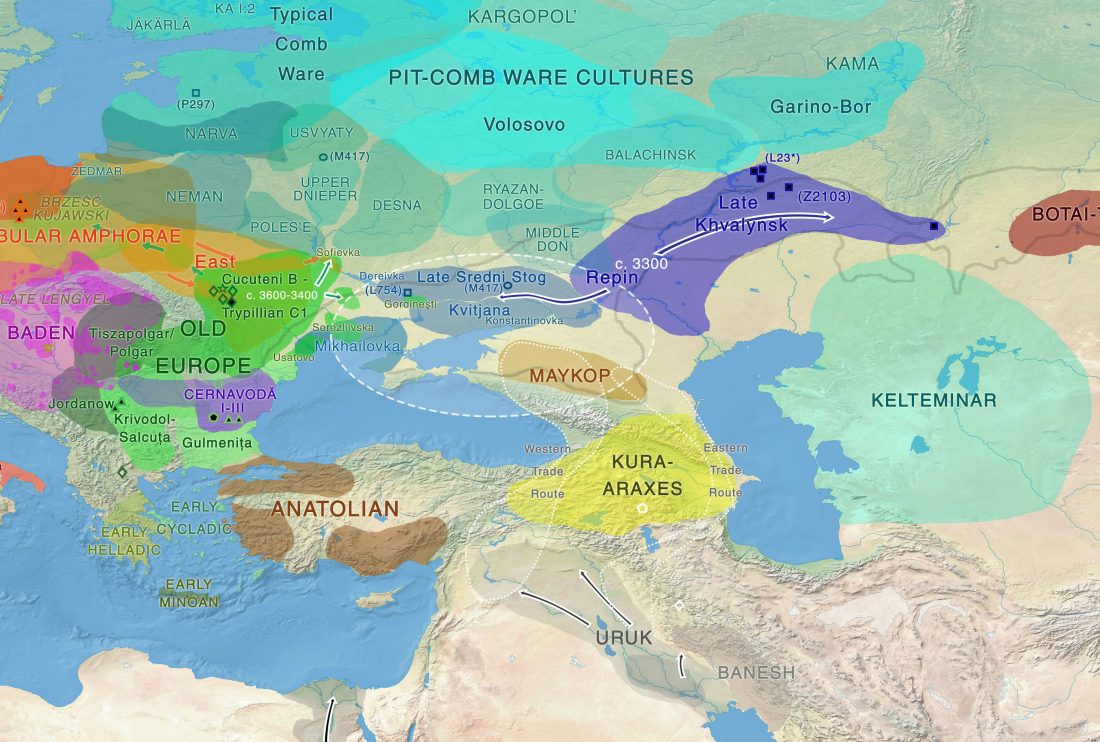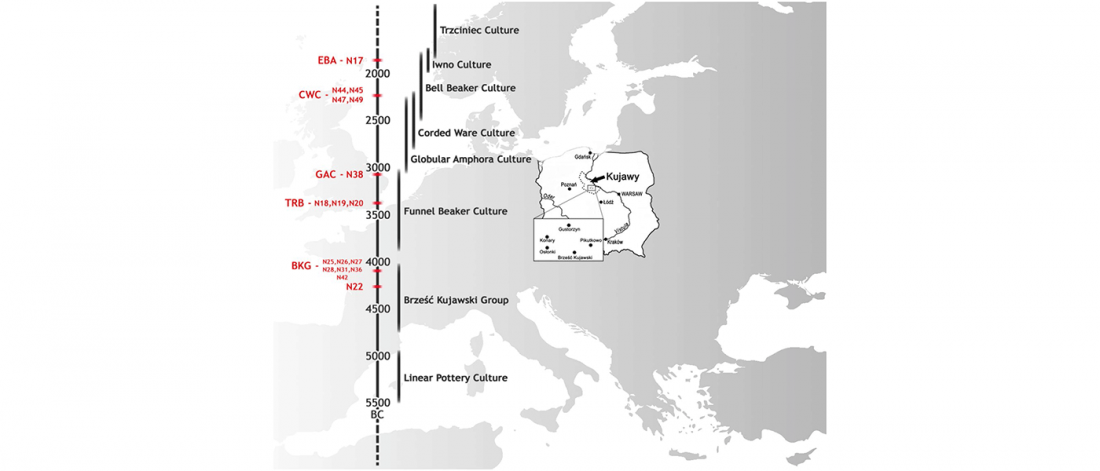
Open access A genomic Neolithic time transect of hunter-farmer admixture in central Poland, by Fernandes et al. Scientific Reports (2018).
Interesting excerpts (emphasis mine, stylistic changes):
… Read the rest “Resurge of local populations in the final Corded Ware culture period from Poland”Most mtDNA lineages found are characteristic of the early Neolithic farmers in south-eastern and central Europe of the Starčevo-Kőrös-Criş and LBK cultures. Haplogroups N1a, T2, J, K, and V, which are found in the Neolithic BKG, TRB, GAC and Early Bronze Age samples, are part of the mitochondrial ‘Neolithic package’ (which also includes haplogroups HV, V, and W) that was introduced to Europe with farmers migrating from Anatolia at the onset of the