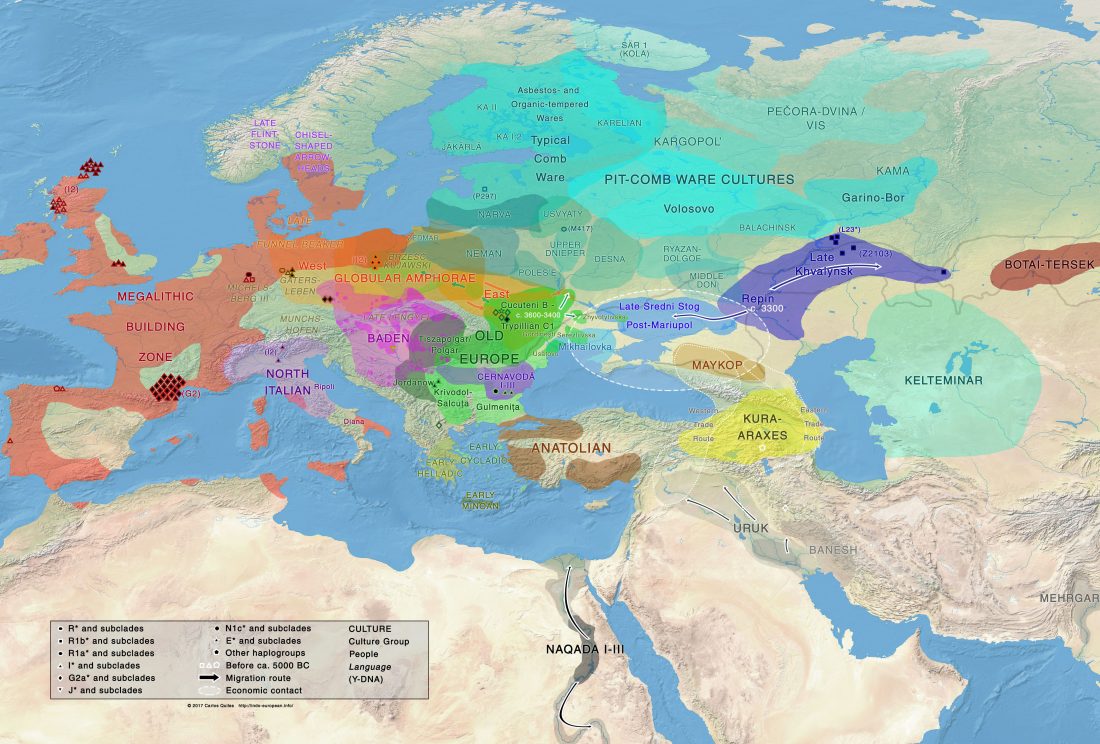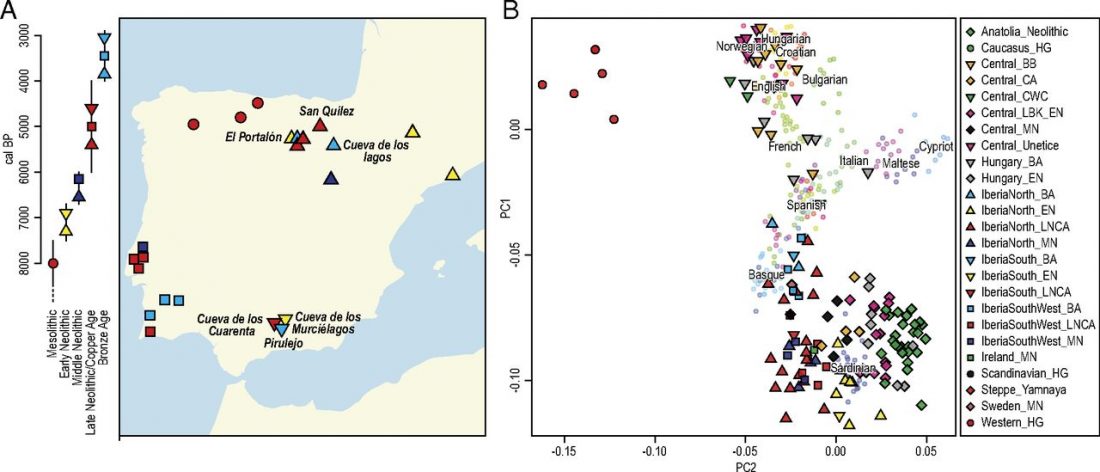
New open access paper Four millennia of Iberian biomolecular prehistory illustrate the impact of prehistoric migrations at the far end of Eurasia, by Valdiosera, Günther, Vera-Rodríguez, et al. PNAS (2018) published ahead of print.
Abstract (emphasis mine)
… Read the rest “Iberian prehistoric migrations in Genomics from Neolithic, Chalcolithic, and Bronze Age”Population genomic studies of ancient human remains have shown how modern-day European population structure has been shaped by a number of prehistoric migrations. The Neolithization of Europe has been associated with large-scale migrations from Anatolia, which was followed by migrations of herders from the Pontic steppe at the onset of the Bronze Age. Southwestern Europe was one of the last parts of