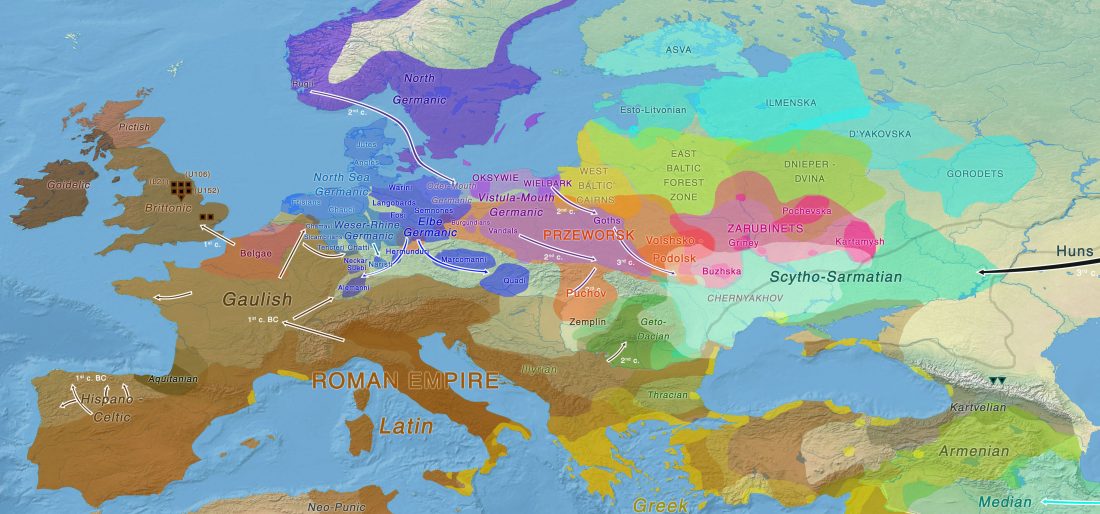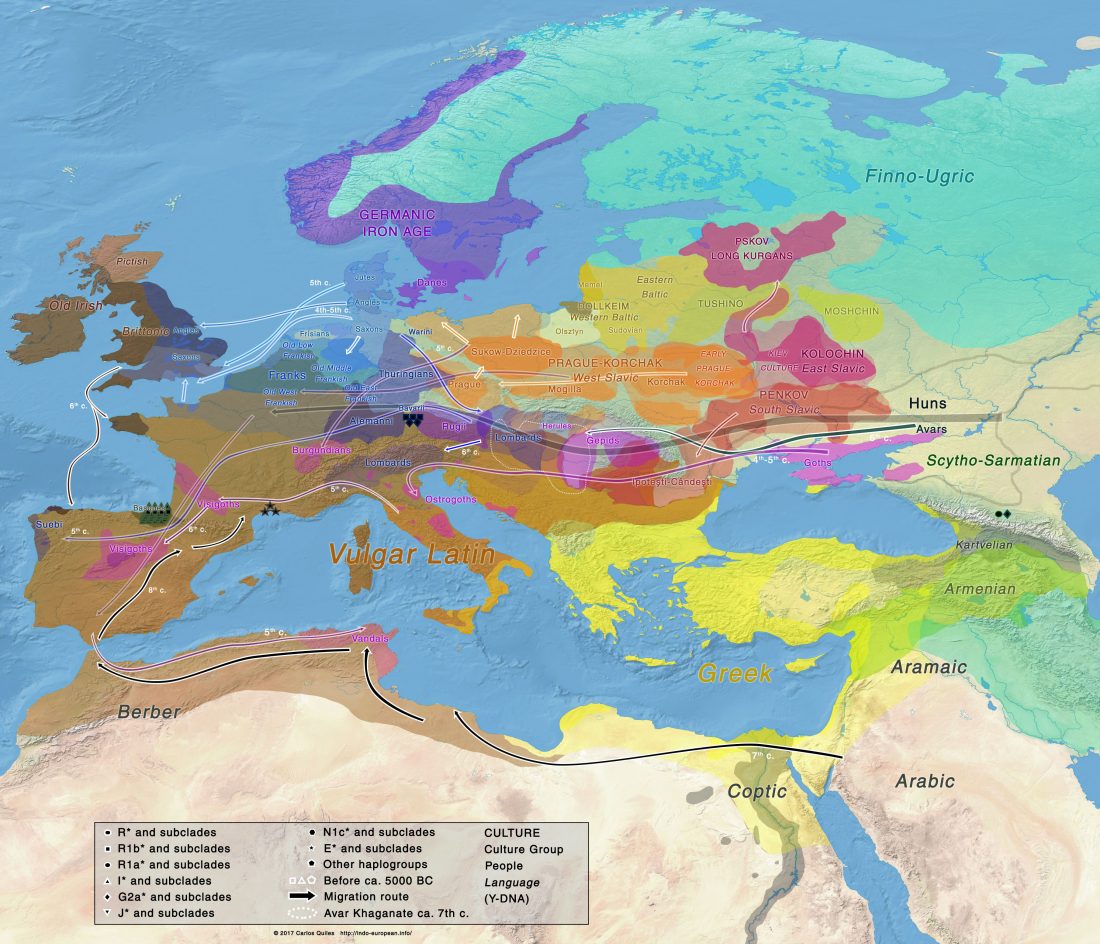
New open access paper Population genomic analysis of elongated skulls reveals extensive female-biased immigration in Early Medieval Bavaria, by Veeramah, Rott, Groß, et al. PNAS (2018), published ahead of print.
First, a bit of context on the Bavarii:
… Read the rest “Genomic analysis of Germanic tribes from Bavaria show North-Central European ancestry”Europe experienced a profound cultural transformation between Late Antiquity and the Middle Ages that laid the foundations of the modern political, social, and religious landscape. During this period, colloquially known as the “Migration Period,” the Roman Empire gradually dissolved, with 5th and 6th century historiographers and contemporary witnesses describing the formation and migration of numerous Germanic peoples, such as the Goths,