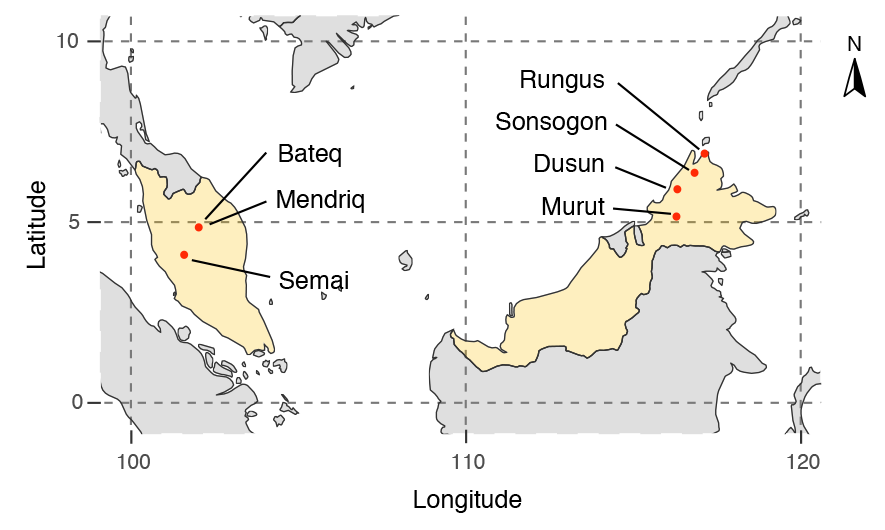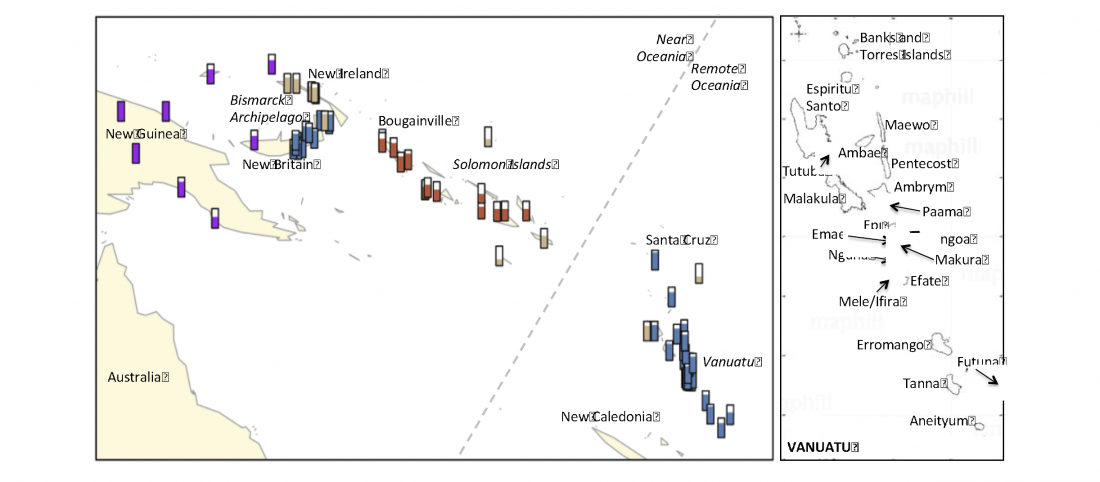
Interesting preprint at BioRxiv by the team of the Reich lab, Population Turnover in Remote Oceania Shortly After Initial Settlement, by Mark Lipson, Pontus Skoglund, Matthew Spriggs, et al. (2018).
Abstract (emphasis mine):
… Read the rest “Population turnover in remote Oceania shortly after initial settlement”Ancient DNA analysis of three individuals dated to ~3000 years before present (BP) from Vanuatu and one ~2600 BP individual from Tonga has revealed that the first inhabitants of Remote Oceania (“First Remote Oceanians”) were almost entirely of East Asian ancestry, and thus their ancestors passed New Guinea, the Bismarck Archipelago, and the Solomon Islands with minimal admixture with the Papuan groups they encountered. However, all