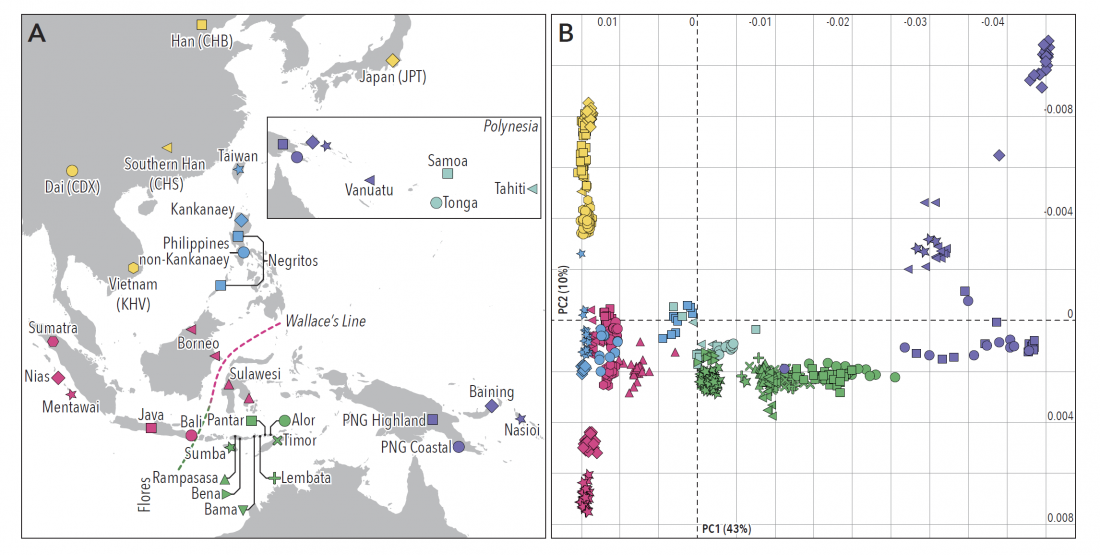
An open access article Complex patterns of admixture across the Indonesian archipelago, by Hudjashov et al. (2017), has appeared in Molecular Biology and Evolution, and clarifies further the Austronesian (AN) expansion.
Abstract:
… Read the rest “Islands across the Indonesian archipelago show complex patterns of admixture”Indonesia, an island nation as large as continental Europe, hosts a sizeable proportion of global human diversity, yet remains surprisingly under-characterized genetically. Here, we substantially expand on existing studies by reporting genome-scale data for nearly 500 individuals from 25 populations in Island Southeast Asia, New Guinea and Oceania, notably including previously unsampled islands across the Indonesian archipelago. We use high-resolution analyses of haplotype diversity to reveal fine