Öffentlich zugängliche Tracking Five Millennia of Horse Management with Extensive Ancient Genome Time Series, von Fages et al. Cell (2019).
Interessante Auszüge:
Der früheste archäologische Beweis für das Melken, Aufspannen und Korrallieren von Pferden findet sich in der ∼5.500 Jahre alten Botai-Kultur der zentralasiatischen Steppen (Gaunitz et al., 2018, Outram et al., 2009; siehe Kosintsev und Kuznetsov, 2013 für Diskussion). Botai-ähnliche Pferde sind jedoch nicht die direkten Vorfahren moderner Domestizierter, sondern der Pferde von Przewalski (Gaunitz et al., 2018). Die genetische Herkunft moderner Domestizierter Pferde ist daher weiterhin umstritten. Vorgeschlagene Kandidaten sind die pontisch-kaspischen Steppen (Anthony, 2007), Anatolien (Arbuckle, 2012, Benecke, 2006) und Iberia (Uerpmann, 1990, Warmuth et al., 2011). . Unabhängig vom Ursprung der Domestikation ist bekannt, dass sich das Pferdegenom in den letzten 2300 Jahren signifikant verändert hat (Librado et al., 2017, Wallner et al., 2017, Wutke et al., 2018). Wann und in welchem Kontext solche Veränderungen stattfanden, ist jedoch weitgehend unbekannt.
Um die Herkunft von Hauspferden zu klären und ihre spätere Transformation durch vergangene Reitzivilisationen aufzudecken, haben wir DNA-Daten aus 278 Subfossilien von Pferden generiert, deren Alter sich größtenteils über die letzten sechs Jahrtausende erstreckt (n = 265, 95%) (Abbildungen 1A und 1B; Tabelle S1; STAR-Methoden). Der endogene DNA-Gehalt war mit der wirtschaftlichen Sequenzierung von 87 neuen Pferdegenomen bis zu einer durchschnittlichen Bedeckungstiefe von 1,0- bis 9,3-fach (Median = 3,3-fach; Tabelle S2) vereinbar.Dies verdoppelt die Anzahl der bisher charakterisierten Genome von Pferden um mehr als das Doppelte. Mit insgesamt 129 alten Genomen, 30 modernen Genomen und neuen Daten im Genommaßstab von 132 alten Personen (0,01- bis 0,9-fach, Median = 0,08-fach) stellt unser Datensatz die größte Zeitreihe im Genommaßstab dar, die für a veröffentlicht wurde nicht-menschlicher Organismus (Tabellen S2, S3 und S4; STAR-Methoden).
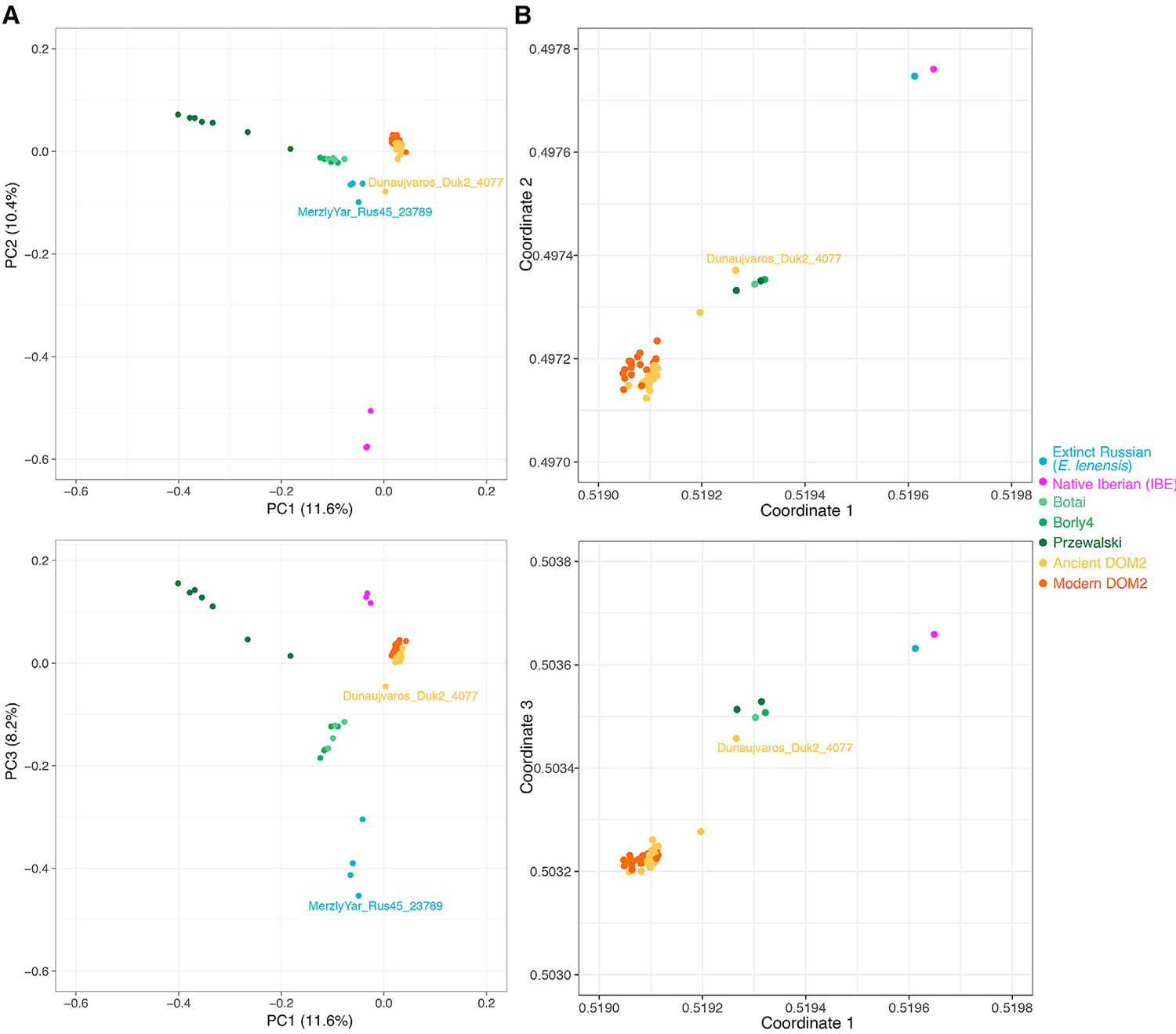
(B) Visualisierung der genetischen Affinitäten zwischen Individuen, wie durch den struct-f4-Algorithmus und 878.475 f4-Permutationen gezeigt. Die f4-Berechnung wurde an Nukleotidtransversionen konditioniert, die in allen Gruppen vorhanden waren, wobei die Proben wie in TreeMix-Analysen gruppiert wurden (Abbildung 3). Im Gegensatz zu PCA messen f4-Permutationen die genetische Drift entlang innerer Äste. Sie zeigen daher eher die Substruktur der alten Bevölkerung.
Entdeckung zweier unterschiedlicher und ausgestorbener Abstammungslinien von Pferden
Hauspferde und Przewalski Pferde sind die einzigen zwei noch existierenden Pferdestämme (Der Sarkissian et al., 2015). Eine andere Abstammungslinie wurde genetisch aus drei Knochen identifiziert, die vor 43.000–5.000 Jahren datiert waren (Librado et al., 2015, Schubert et al., 2014a). Es zeigte morphologische Affinitäten zu einer ausgestorbenen Pferdeart, die als Equus lenensis beschrieben wird (Boeskorov et al., 2018). Wir stellen jetzt fest, dass sich diese ausgestorbene Linie nach der Hauptkomponentenanalyse (PCA), der phylogenetischen und der f3-Outgroup-Clusterbildung einer 24.000 Jahre alten Probe aus der Tuva-Republik innerhalb dieser Gruppe auch auf Südsibirien ausdehnte (Abbildungen 3, 5A) und S7A). Diese neue Probe (MerzlyYar_Rus45_23789) trägt eine extrem divergierende mtDNA, die nur auf den Neusibirischen Inseln vor 33.200 Jahren (Orlando et al., 2013) gefunden wurde (Abbildung 6A; STAR-Methoden) und in den drei zuvor sequenzierten Knochen nicht vorhanden ist. Dies deutet darauf hin, dass eine unterschiedliche Geisterlinie von Pferden zur genetischen Abstammung von MerzlyYar_Rus45_23789 beigetragen hat. Sowohl der Zeitpunkt als auch der Ort des genetischen Kontakts zwischen E. lenensis und dieser Geisterlinie bleiben jedoch unbekannt.
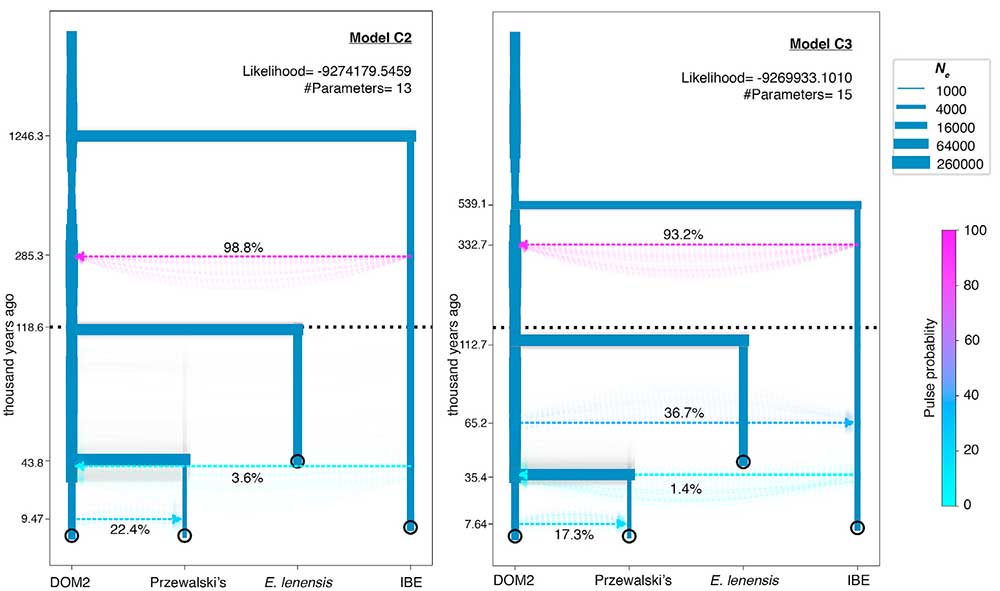
Modellierung der Demographie und Vermischung ausgestorbener und erhaltener Pferdestämme
Phylogenetische Rekonstruktionen ohne Genfluss zeigten, dass IBE vor der Divergenz zwischen DOM2 und Przewalski-Pferden differenzierte (Abbildung 3; STAR-Methoden). Die Berücksichtigung einer Migrationskante in TreeMix deutete jedoch auf eine engere Verwandtschaft mit einer einzelnen ungarischen DOM2-Probe aus der 3. Mühle hin. BCE (Dunaujvaros_Duk2_4077), mit umfangreichem genetischen Beitrag (38,6%) vom Zweig der Vorfahren zu allen Pferden (Abbildung S7B). Dies und das extrem divergente IBE Y-Chromosom (Abbildung 6B) lassen vermuten, dass eine divergente, aber noch nicht identifizierte Geisterpopulation zum Erbgut von IBE beigetragen haben könnte.
Ablehnung des iberischen Beitrags zu modernen Haushalten
Die hier charakterisierten Genomsequenzen von vier 4.800- bis 3.900-jährigen IBE-Exemplaren ermöglichten es uns, die laufenden Debatten über den möglichen Beitrag von Iberia zur Domestikation von Pferden zu klären (Benecke, 2006, Uerpmann, 1990, Warmuth et al., 2011). Die Berechnung des sogenannten fG-Quotienten (Martin et al., 2015) lieferte eine minimale Grenze für den IBE-Beitrag an DOM2-Mitglieder (Cahill et al., 2013) (Abbildung 7A). Das Maximum einer solchen Schätzung wurde in der ungarischen Probe Dunaujvaros_Duk2_4077 gefunden (~ 11,7% –12,2%), was mit der TreeMix-Clusterung mit IBE übereinstimmt, wenn eine Migrationskante berücksichtigt wird (Abbildung S7B). Es wurde zuvor vorgeschlagen, dass dieses Exemplar einen Vorfahren mit einer noch nicht identifizierten Population teilt (Gaunitz et al., 2018). Die Berechnung der f4-Statistik zeigt, dass diese Population nicht mit E. lenensis, sondern mit IBE verwandt ist (7B; STAR-Methoden). Daher haben IBE oder Pferde, die eng mit IBE verwandt sind, die Abstammung von Tieren beigetragen, die in einem Handelszentrum der frühen Bronzezeit in Ungarn ab der späten 3. Mühle gefunden wurden. BCE. Dies könnte darauf hindeuten, dass während des Bell-Beaker-Phänomens ein Fernaustausch von Pferden stattfand (Olalde et al., 2018). Die fG- Minimalgrenze für den IBE-Beitrag zu einem eisenzeitlichen spanischen Pferd ( ElsVilars_UE4618_2672 ) war weiterhin wichtig (~ 9,6% –10,1%), was darauf hindeutet, dass ein genetischer IBE-Einfluss in Iberia bis mindestens zum 7. Jahrhundert v . Die fG-Schätzungen waren jedoch für fast alle untersuchten alten und modernen Pferde begrenzter (Median = ~ 4,9% –5,4%; Abbildung 7A).
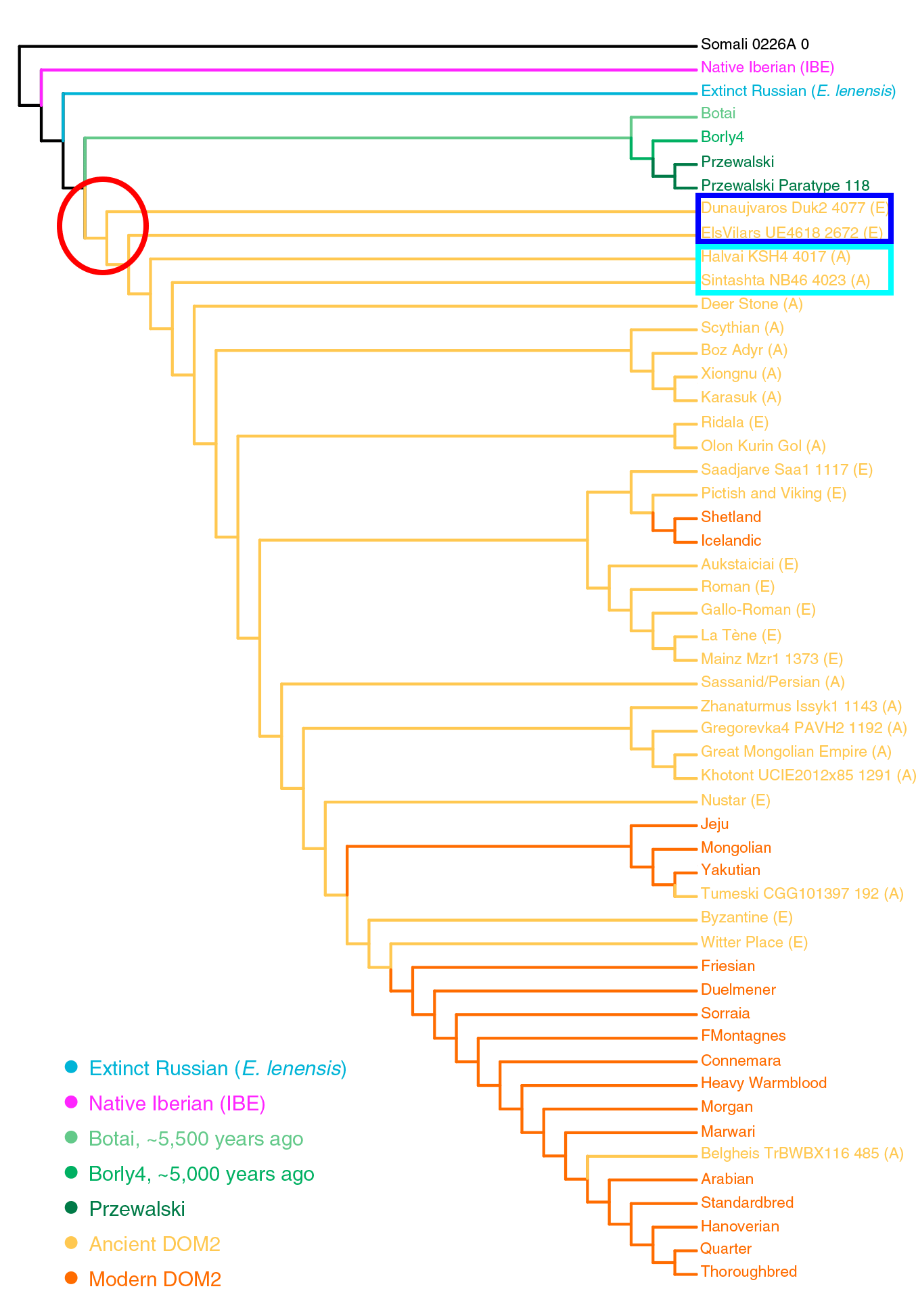
Pferde aus der Eisenzeit
Die Y-Chromosomennukleotid-Diversität (π) nahm in beiden Kontinenten in den letzten 2000 Jahren stetig ab, fiel jedoch erst nach 850–1.350 CE auf das heutige Niveau (Abbildungen 2B und S2E; STAR-Methoden). Dies steht im Einklang mit der Dominanz einer 1000 bis 700 Jahre alten orientalischen Haplogruppe in den meisten modernen Gestüten (Felkel et al., 2018, Wallner et al., 2017). Unsere Daten zeigen auch, dass der wachsende Einfluss spezifischer Hengstlinien nach der Renaissance (Wallner et al., 2017) für einen 3,8- bis 10,0-fachen Rückgang der Y-Chromosomendiversität verantwortlich war.
Anschließend berechneten wir die Y-Chromosomen-π-Schätzungen in früheren Kulturen, die von mindestens drei Männern repräsentiert wurden, um die historischen Zusammenhänge zu klären, die die Y-Chromosomen-Diversität am stärksten beeinflussten. Dies bestätigte den oben beobachteten zeitlichen Verlauf, da byzantinische Pferde (287–861 n. Chr.) Und Pferde aus dem großen mongolischen Reich (1.206–1.368 n. Chr.) Eine begrenzte, jedoch übermoderne Vielfalt aufwiesen. Bronzezeitliche Hirschsteinpferde aus der Mongolei, mittelalterliche Aukštaičiai-Pferde aus Litauen (9.-10. Jh. [9.-10. Jh.]) Und eisenzeitliche Pazyryk-Skythenpferde wiesen ähnliche Unterschiede auf (0,000256–0,000267) (Abbildung 2A). Die Diversität war jedoch bei Pferden von La Tène, Roman und Gallo-Roman größer , bei denen das Verhältnis von Y zu autosomalem π nahe bei 0,25 lag. Dies steht im Gegensatz zu modernen Pferden, bei denen eine ausgeprägte Auswahl spezifischer Patrilinien das Verhältnis von Y zu autosomalem π deutlich unter 0,25 (0,0193–0,0396) treibt (Abbildung 2A). Die in La Tène, Roman und Gallo-Roman Pferden gefundenen π-Verhältnisse von nahezu 0,25 zu autosomal deuten auf Zuchtstrategien hin, die einen gleichmäßigen Fortpflanzungserfolg bei Hengsten oder einen gleich voreingenommenen Fortpflanzungserfolg bei beiden Geschlechtern beinhalten (Wilson Sayres et al., 2014).
Die Linie wird in diesem Artikel von einer bestimmten Abstammung definiert, genauso in vielen anderen Artikeln der Genetik verwendet wird. Ich behalte diese Nomenklatur unten. Es sollte nicht mit den “Linien” verwechselt werden, die sich auf Y-Chromosomen (oder mtDNA)-Haplogruppen beziehen.
Unterstützend für die „archaische“ Natur der ungarischen GBK-Pferde, die aus der pontisch-kaspischen Steppe heranwachsen, sind:
- Unter den Y-Chromosomenlinien ist die gemeinsame Gruppe von Botai-Borly4 (eng verwandt mit DOM2) , skythischen Pferden aus Aldy Bel (Arzhani), eisenzeitlichen Pferden aus Estland (Ridala), Pferden aus der Xiongnu-Kultur (Uushgiin Uvur) und Römische Pferde aus Autricum (Chartres).
- Unter den mtDNA-Linien zeigt sich die gemeinsame Gruppe aus Botai-Proben, LebyazhinkaIV NB35 und verschiedenen eurasischen Domestizierten, einschließlich vieler alter westeuropäischer, eine wahrscheinliche Ausweitung bestimmter Subklassen im Osten und Westen mit der Repin-Kultur .
(…) DOM2 steuerte 22% zu Przewalskis Vorfahren bei. 9,47 kya, was eher auf das holozäne Optimum als auf die eneolithische Botai-Kultur (∼5,5 kya) als auf eine Periode des Bevölkerungskontakts hindeutet. Diese Introgression vor Botai könnte die Y-Chromosomentopologie erklären, bei der Botai-Pferde Berichten zufolge zwei verschiedene segregierende Haplogruppen tragen: Eine befand sich in der Phylogenie in einer basalen Position, während die andere eng mit DOM2 verwandt war . Es ist jedoch bekannt, dass mehrere Beimischungspulse entlang der Divergenz von DOM2 und der Botai-Borly4-Linie aufgetreten sind, einschließlich eines 2,3%igen Beitrags nach Borly4 zu DOM2 und eines jüngeren 6,8%igen DOM2-Angriffs auf Przewalski-Pferde (Gaunitz et al., 2018). Die Parameter des Modells C2 berücksichtigen all diese Faktoren als einen einzigen Zumischungsimpuls und bilden wahrscheinlich den Durchschnitt der Beiträge all dieser multiplen Ereignisse.
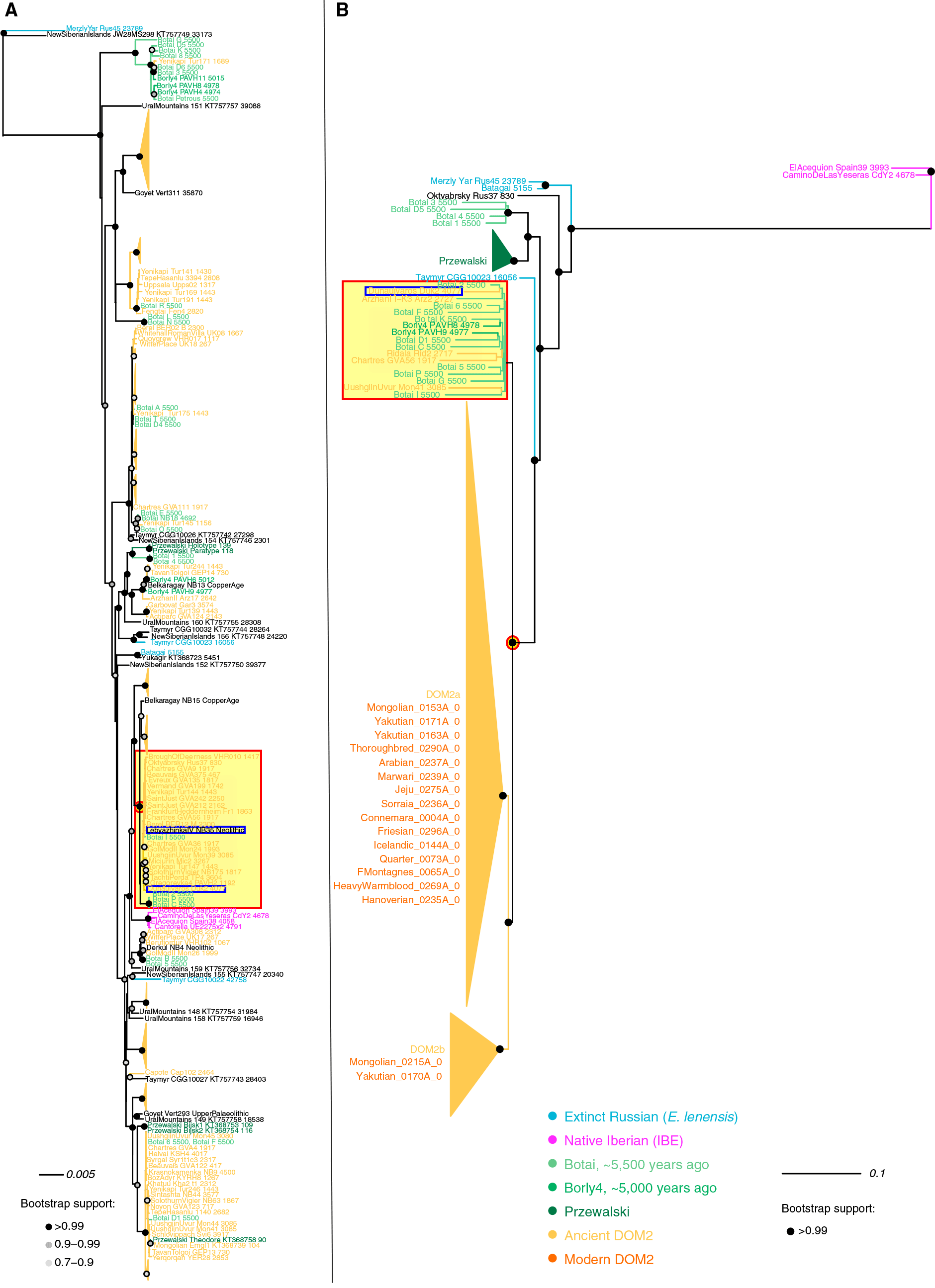
B) Bester Y-Chromosomen-Maximum-Likelihood-Baum (GTRGAMMA-Substitutionsmodell) ohne Outgroup. Knotenunterstützung wird als Bruchteil von 100 Bootstrap-Pseudo-Replikaten angegeben. Bootstrap-Unterstützungen, die unter 90% liegen, werden nicht angezeigt. Die Wurzel wurde auf den Baummittelpunkt gelegt. Siehe auch Tabelle S5 für Datensatzinformationen.
Das Bild wurde aus dem Papier modifiziert, einschließlich eines roten Quadrats in archaischen Gruppen, das die ungarische Stichprobe enthält, und eines roten Kreises um den wahrscheinlich häufigsten Ahnenhengst und die Stute aus der pontisch-kaspischen Steppe.
Der Artikel kann kein detailliertes Bild der Domestizierung alter Pferde bieten, aber es ist ein weiterer Schritt, um zu zeigen, dass Repin / Jamnaja die wahrscheinlichste Quelle für die Ausbreitung des Hauspferdes von eurasischen domestizierten Pferden ist. Noch interessanter ist, dass Jamnaja-Siedler in Ungarn wahrscheinlich eine uralte Linie dieses Pferdes verbreiteten, während sie sich gleichzeitig mit der Kultur des klassischen Glockenbechers verbreiteten. Bemerkenswerte Parallelen finden sich also zwischen:
- die Abstammungslinie der Dunaujvarus Duk2 4077-Probe und die Ausbreitung des Nordwestindogermanisches (ca. 3000-2500 v. Chr.).
- die Abstammungslinie der Halvai-Proben KSH4 4017 und Sintashta NB46 4023 und die spätere Ausbreitung des Urindoiranisches (ca. 2500-2000 v. Chr.).
Die Ausweitung einer alten Linie von Pferdedomizilen, die mit Jamnaja von Ungarn / östliche Glockenbechers verwandt sind, scheint durch die vor-iberische Pferde-Probe von Vilars I, Els Vilars4618 2672 (ca. 700-550 v. Chr.), wahrscheinlich iberischer GBK Abstammung, bestätigt zu werden, als es sich von eine Linie handelt, die älter ist als die indoiranischen, die nur später die meisten europäischen Linien ersetzte.
ANMERKUNG. Informationen zu bekannten Kontakten zwischen Jamnaja und Ur-Glockenbechers kurz vor der Ausbreitung Glockenbecherleuten finden Sie in einem kürzlich veröffentlichten Beitrag über Spitze Jamnaja-Gruppen .
Die Ergebnisse des Artikels bestätigen die Ausbreitung des Hauspferdes zuerst (und hauptsächlich) durch das Steppenbiom , wobei die Aubreitung der Urindogermaner zuerst nachgeahmt und dann schrittweise (oder nicht so schrittweise) durch Linien ersetzt wird, die während der Verbreitung der Reitsteppenkulturen nach Westen ins Europa gebracht wurden, während der Bronzezeit, Eisenzeit und Mittelalter. Die Erweiterung korreliert auch gut mit der bekannten Ausbreitung der Tierhaltung und des Pastoralismus vor 2000 v.C.:
BEARBEITUNG (3. MAI 2019): Eine aktuelle Erinnerung an diese parallelen Entwicklungen von David Reich in Einsichten in Spracherweiterungen aus der alten DNA :
- Die Ausbreitung der Jamnaja nach Westen “mit Pferden und Wagen”, mit einer homogeneren Abstammung in modernen Europäern aufgrund späterer Wanderungen aus dem Osten (und Norden):
DR: inference is that two major migrations: farmers from Anatolia, followed by steppe pastoralists. Who are they? They took horses and wagons and spread. See rapid 90% pop turn over in Britain. Similar timing in Iberia, but a bit less turnover, and more period of overlap
— Joshua G. Schraiber? (@jgschraiber) May 2, 2019
- “Nachkommen” von Jamnaja (nachdem die Kultur bereits “tot” war), die sich hauptsächlich mit Schnurkeramik-Vorfahren nach Osten ausdehnten:
DR: spread of steppe ancestry to the east likely a result of spread of yamnaya descendents, since yamnaya were dead. High genetic similarity to corded ware people.
— Joshua G. Schraiber? (@jgschraiber) May 2, 2019
Ein weiterer kürzlich veröffentlichter öffentlich zugänglicher Artikel zur Domestikation der Pferde ist The horse Y chromosome as an informative marker for tracing sire lines, von Felkel et al. Scientific Reports (2019).
Siehe auch
- The genetic and cultural barrier of the Pontic-Caspian steppe – forest-steppe ecotone
- Mitogenomes suggest rapid expansion of domesticated horse before 3500 BC
- Origin of horse domestication likely on the North Caspian steppes
- Ancient DNA upends the horse family tree
- Domesticated horse population structure, selection, and mtDNA geographic patterns
- About Scepters, Horses, and War: on Khvalynsk migrants in the Caucasus and the Danube
- Steppe and Caucasus Eneolithic: the new keystones of the EHG-CHG-ANE ancestry in steppe groups
- Domestication spread probably via the North Pontic steppe to Khvalynsk… but not horse riding
- Differing modes of animal exploitation in North Pontic Eneolithic and Bronze Age Societies